RESUMEN: En Ecuador existen bosques montanos bajos de gran importancia biológica que proveen de servicios ecosistémicos como el almacenamiento de carbono. Por esta razón, se desarrolló una investigación que permitió elaborar modelos alométricos mediante la cuantificación de carbono en el bosque montano bajo. Se estableció una parcela permanente de una hectárea y se midieron todos los árboles con diámetro a la altura del pecho (DAP1,30m) ≥ 5 cm. Las variables que se usaron para la generación de los modelos fueron DAP y altura total (ht). La biomasa arbórea fue de 322,37 t ha-1 y el carbono total estimado fue de 161,07 t ha-1. En los modelos alométricos del estrato arbóreo, las variables que mejor se ajustaron fueron DAP, DAP2 y ht. La familia botánica con mayor biomasa (69,95 t ha-1) y carbono (34,98 t C ha-1) fue Moraceae, que conjuntamente con Lauraceae, Myristicaceae y Rubiaceae almacenan el 57% de carbono (91,55 t C ha-1) del bosque montano bajo. De las 179 especies botánicas registradas, las especies Pseudolmedia laevigata, Nectandra reticulata, Otoba parvifolia, Vochysia paraensis, Ficus sp. y Elaeagia obovata almacenan el 51% (81 t C ha-1) del carbono total estimado.
Palabras claves: biomasabiomasa,servicio ecosistémicoservicio ecosistémico,LauraceaeLauraceae.
ABSTRACT: In Ecuador, there are montane forests of great biological importance that provide ecosystem services such as carbon storage. For this reason, we did this research for generate allometric models through by quantifying carbon in lower montane forests. We established one permanent plot of one hectare and measured all the trees with diameter at breast height (DBH1,30m) ≥ 5 cm. The variables that were used to generate the models were: DBH and total height (th). The tree biomass was 322.37 t ha-1 and the estimated total carbon was 161.07 t ha-1. In the allometric models of the arboreal stratum, the variables that were adjusted best were: DBH, DBH2 and th. The botanical family with more biomass (69.95 t ha-1) and carbon (34.98 t C ha-1) was Moraceae, which together with Lauraceae, Rubiaceae and Myristicaceae stored the 57% carbon (91.55 t C ha-1) in the lower montane forest. Of the 179 botanical species registered, Pseudolmedia laevigata, Nectandra reticulata, Otoba parvifolia, Vochysia paraensis, Ficus sp and Elaeagia obovata stored 51% (81 t C ha-1) of carbon total estimated.
Keywords: biomass, ecosystem service, Lauraceae.
Nota Técnica
MODELOS ALOMÉTRICOS PARA ESTIMAR EL ALMACENAMIENTO DE CARBONO DE BOSQUES MONTANOS BAJOS EN EL SUR DEL ECUADOR
ALLOMETRIC MODELS TO ESTIMATE STOCKS CARBON IN THE LOWER MONTANE FORESTS IN SOUTH OF ECUADOR
Universidade Federal de Santa Maria
Recepción: 10 Mayo 2016
Aprobación: 28 Julio 2017
Los bosques son ecosistemas que capturan y conservan el carbono mediante la fijación y almacenamiento a través de la fotosíntesis (RÜGNITZ; CHACÓN; PORRO, 2009). La cantidad de carbono encima y abajo del suelo varía en función de variables bioclimáticas, de la composición florística, la edad y la densidad de población de cada estrato por comunidad vegetal, tasa de crecimiento, longevidad, ubicación, clima y rotación (ZAMBRANO; FRANQUIS; INFANTE, 2004; FONSECA; ALICE; REY, 2009; VALDÉS RAMÍREZ, 2012; RODRÍGUEZ; CARGUA, 2013; ARÉVALO, 2015).
La cuantificación de la biomasa en base al crecimiento de la vegetación en los ecosistemas forestales es importante para estimar el carbono almacenado y las tasas anuales de fijación, un tema relevante por sus implicaciones directas con el calentamiento global (CIFUENTES JARA, 2010; VÁSQUEZ; ARELLANO, 2012). Se ha demostrado que los ecosistemas forestales, con manejo adecuado, pueden secuestrar más carbono que otros ecosistemas terrestres, contribuyendo a la mitigación del cambio climático mediante la reducción de la tasa de aumento de CO2 en la atmósfera y estabilización de las concentraciones atmosféricas (BENNACEUR et al., 2005; YEPES QUINTERO et al., 2011).
Para estimar la cantidad de carbono en la biomasa de un bosque existen métodos directos e indirectos (BROW, 1997; BROW; CABARLE; LIVERNASH, 1997; SCHLEGEL; GAYOSO; GUERRA, 2001). Entre los métodos indirectos están las ecuaciones o modelos alométricos basados en análisis de regresión lineal o no lineal. Estos utilizan variables medidas en el campo denominadas regresoras o independientes tales como: diámetro a la altura del pecho (DAP), altura comercial (hc) y altura total (ht), incremento diamétrico, área basal y densidad específica de la madera (CHAVE et al., 2005; JADÁN; TORRES; GÜNTER, 2012).
En el Ecuador, según el Ministerio del Ambiente (MAE) (ECUADOR, 2012a), existen bosques de tierras bajas de gran importancia biológica con una cobertura vegetal significativa para la conservación y provisión de servicios ecosistémicos como el almacenamiento de carbono, el cual, tendría muchas implicaciones negativas en el momento de ser liberado a la atmósfera por actividades antrópicas. Uno de estos ecosistemas es el bosque montano bajo que forma parte de la cuenca del río Amazonas (un verdadero sumidero de carbono). Pese a su importancia, este ecosistema forestal se encuentra amenazado, debido a las altas tasas de deforestación, degradación y el avance de la frontera agrícola (LAMBIN; GEIST; LEPERS, 2003; MALHI; PHILLIPS, 2004; ORGANIZACIÓN DE LAS NACIONES UNIDAS PARA LA AGRICULTURA Y LA ALIMENTACIÓN, 2007b; BACCINI et al., 2012). Por ejemplo, la deforestación promedio en Ecuador para el período 2000 - 2008 fue de 77.647 ha año-1 mientras que para el período 2008 - 2012 fue de 65.880 ha año-1 (ECUADOR, 2014), específicamente en los bosques amazónicos la deforestación promedio para el periodo de 2000 - 2008 fue de 16.430 ha año-1 (ECUADOR, 2012b). Por lo tanto la caracterización y valoración de sus servicios ecosistémicos, como el carbono almacenado, es un tema relevante para la política nacional de cambio climático (ECUADOR, 2012a), así como, para disminuir las tasas de deforestación de estos ecosistemas. Los objetivos de esta investigación fueron: cuantificar la biomasa almacenada encima del suelo, ajustar modelos alométricos para estimar la biomasa y determinar las familias y especies botánicas que acumulan la mayor cantidad de carbono, en el bosque montano bajo del Sur de Ecuador.
El estudio se realizó en el bosque montano bajo de la microcuenca “El Padmi” de la Universidad Nacional de Loja, ubicado en el corredor fluvial del Río Zamora, entre 775 - 1150 m de altitud, con precipitación media anual de 1978 mm año-1, clima semicálido húmedo, temperatura media anual de 23 ºC y suelos jóvenes del grupo entisoles e inceptisoles, en la clasificación de la FAO este orden de suelos entra típicamente en el Grupo de Cambisoles (SIERRA et al., 1999; CERÓN et al., 2003; VALAREZO, 2004; ORGANIZACIÓN DE LAS NACIONES UNIDAS PARA LA AGRICULTURA Y LA ALIMENTACIÓN, 2007a). Según Holdridge (1967), el área de estudio pertenece a la zona de vida bosque muy húmedo tropical (Figura 1).
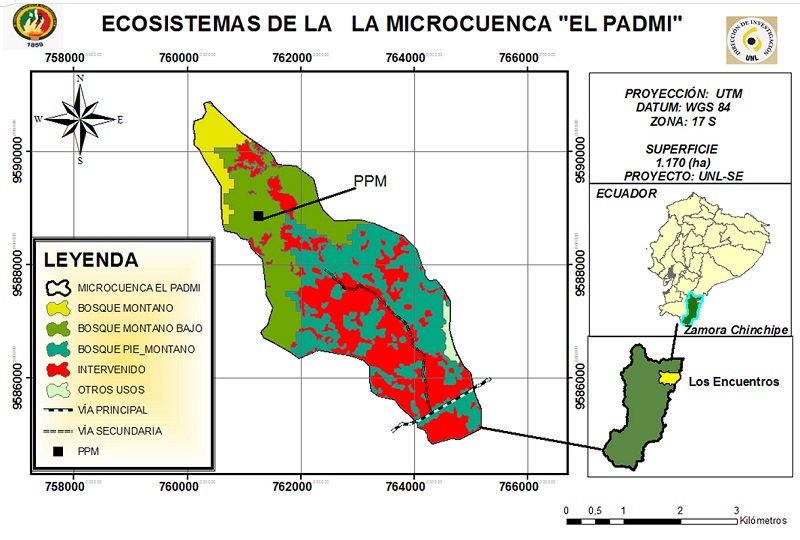
Figura 1
Ubicación geográfica del área de estudio, microcuenca “El Padmi, Zamora Chinchipe, sur de Ecuador
Se instaló una parcela permanente considerando la topografía del terreno y la heterogeneidad del bosque. La parcela permanente de 1 ha fue subdivida en 25 unidades de muestreo de 400 m2, donde se inventarió los individuos ≥ 5 cm de DAP, para obtener el volumen total.
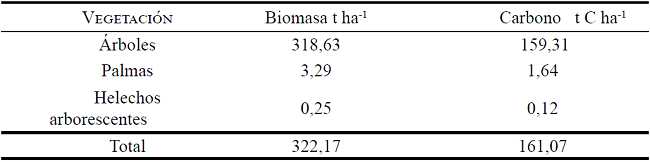
Para estimar el carbono almacenado en la biomasa de los árboles, palmas y helechos arborescentes del bosque se aplicó la metodología propuesta por Ravindranath y Ostwald (2008), quienes indican que la biomasa se obtiene del volumen de cada árbol multiplicándolo por la densidad de la madera (ecuación 1), esta última variable fue obtenida de lista de densidades de maderas de la FAO (ZANNE et al., 2009). Para obtener el carbono fijado por árbol (ecuación 2) la biomasa se multiplica por el factor de conversión 0,5 propuesto por el Grupo Intergubernamental de Expertos sobre el Cambio Climático (IPCC) (PENMAN et al., 2003). Los valores obtenidos fueron expresados en toneladas de carbono por hectárea (t C ha-1). El carbono total almacenado se calculó sumando el carbono en cada uno de los compartimentos de almacenamiento (biomasa arbórea, palmas y helechos arborescentes).
Donde: B = biomasa en t ha-1; V = volumen del árbol en m3; = densidad de la madera en m3 kg-1; Cfijado = carbono fijado en t C h-1; f = factor de conversión de biomasa a carbono 0,5 (IPCC, 2006).
Para determinar las variables dendrométricas que influyen sobre la presencia y magnitud de la biomasa se calculó la correlación de Pearson (p<0,05) entre diámetro a la altura del pecho (DAP1.30m), altura total (ht), área basal (G) y volumen total (Vt).
Para construir los modelos se utilizó el programa estadístico Infostat profesional versión 2013 (DI RIENZO et al., 2011). La selección de las ecuaciones alométricas mejor ajustadas se realizó aplicando la metodología propuesta por Segura y Andrade (2008) y Fonseca, Alice y Rey (2009) que proponen los siguientes pasos: los datos que se usen en el análisis de regresión deben cumplir los supuestos de normalidad, independencia y homogeneidad de varianzas. En caso de no cumplirse algunos de ellos, entonces se transforman las variables usando logaritmos (naturales y en base 10), potencias y raíces. Walpole y Myers (1992) indican que una gráfica de valores observados en función de la variable independiente puede indicar la necesidad de transformar los datos a logaritmos. Cuando no sea posible linearizar mediante transformaciones.
Para seleccionar el modelo se consideraron las siguientes variables:
-
Cálculo de indicadores de ajuste: coeficiente de determinación ajustado (R2). Aquellos modelos que presentaron, al menos, un coeficiente no significativo (p > 0,05) respecto a una variable utilizada como independiente o regresora fueron eliminados.
-
Cálculo de indicadores de capacidad predictiva de los modelos: Error Cuadrático Medio Ponderado (ECMP), Criterio de Información Akaike (AIC) y Criterio de información Bayesiano (BIC).
-
Se calificó cada modelo en base a la escala de calificación propuesta por Segura y Andrade (2008). El modelo con menor puntaje en base a la sumatoria de los indicadores fue seleccionado como el mejor, en base a lo siguiente: en el caso del R2 será el de mayor valor y tomará el valor 1, cuando se aproxime a este (escala de 0-1) y en los indicadores de predicción (ECMP; AIC y BIC) los que más se acercan a cero. Los valores de 2, 3, 4 etc., corresponderán al valor del indicador que se aleja de uno (R2) o del cero.
Finalmente, se determinó la familia y especie botánica que almacenan la mayor cantidad de carbono en el bosque montano bajo, para lo cual se realizó un análisis estadístico multivariado (ANAVA) utilizando el programa estadístico Infostat profesional versión 2013 (DI RIENZO et al., 2011).
La biomasa y carbono arbóreo dependen del área basal, altura total y número de individuos de los bosques, como se evidencia en el carbono estimado en el bosque montano bajo, el cual fue de 161,07 t C ha-1 (Tabla 1). Los resultados hallados se encuentran en el rango que señala el EcoSecurities (2007), para los bosques tropicales montanos que están entre 100 y 200 t C ha-1. Así mismo, estos resultados son corroborados por Unger, Homeier y Leuschner (2012) quienes determinaron 344 ± 17 t ha-1 de biomasa respecto al área basal, al transformar la biomasa estimada por Unger, Homeier y Leuschner (2012) a carbono mediante el factor de conversión de 0,5 propuesto por el IPCC tenemos un valor equivalente a 172 ± 8,5 t C ha-1.
Por otro lado, Honorio-Coronado, Vega-Arenas y Corrales-Medina (2015), calcularon un promedio de 193,9 ± 55,6 t ha-1 de biomasa en bosques aluviales de la amazonia peruana, valor diferente al reportado en este estudio para el bosque montano bajo de Ecuador, a pesar de ser ecosistemas con características climáticas similares. En cambio, en estudios realizados en bosques de la amazonia de Brasil, Castilho et al. (2006), determinaron en promedio 327,8 t ha-1 de biomasa y Nogueira (2006) estimo 320,8 t ha-1 de biomasa, valores que concuerdan con los resultados de esta investigación. También, Bertzky et al. (2010) en su estudio sobre carbono, biodiversidad y servicios ecosistémicos realizado en Ecuador, señala que los bosques siempreverdes de tierras bajas de la amazonia almacena mayor cantidad de carbono en relación a ecosistemas como los páramos, lo cual se contrasta con los datos encontrados en la presente investigación.
Sin embargo, Hofstede (1999) menciona que la biomasa de la selva tropical almacena 250 t C ha-1 valor distante al reportado en esta investigación. También, los resultados de esta investigación (bosque montano bajo) difieren con los encontrados por Arévalo (2015), en su investigación realizada en el estrato arbóreo del bosque natural Tinajillas - Limón Indanza en Ecuador, quien estimo 41 t C ha-1. Así mismo, los resultados obtenidos en el bosque montano bajo son diferentes a los reportados por Ponce et al. (2009), en su estudio realizado en la cordillera del Cóndor en Ecuador, cuya estimación de biomasa y carbono fue de 43,7 t ha-1 y 21,8 t C ha-1, respectivamente.
La diferencia en los resultados posiblemente se debe al número de individuos evaluados, que difieren notablemente entre los estudios realizados en la cordillera del Cóndor, bosque natural Tinajillas y el bosque montano bajo. En el bosque montano bajo se evaluaron 1.408 individuos ha-1 y 52 familias botánicas para la cuantificación del almacenamiento de carbono (Tabla 1), en cambio, en el estudio de la cordillera del Cóndor se evaluaron 512,9 individuos ha-1 y 37 familias botánicas, y en el bosque natural Tinajillas - Limón Indanza se evaluaron 925 individuos y 22 familias botánicas.
Biomasa total encima del suelo y reserva de carbono de la vegetación en el bosque montano bajo del sur de Ecuador
Cuando analizamos las correlaciones (Pearson, p < 0,05) entre la biomasa y las variables dasométricas de los árboles constatamos que los resultados estaban fuertemente correlacionados (Tabela 2). Las variables: DAP (m ha-1), área basal (m2 ha-1) y volumen (m3 ha-1) explican significativamente (p < 0,05) la cantidad de carbono encima del suelo debido a que están fuertemente correlacionadas con la biomasa. En cambio, la altura (m) presenta relación media cuya correlación es moderada con la biomasa del bosque montano bajo (Tabla 2). Según, Pearson et al. (2007), las variables de rendimiento dasométricas están altamente relacionadas con la biomasa de un ecosistema, lo que es ratificado con los resultados encontrados en el bosque montano bajo. En cambio, Lal (2005), indica que la biomasa y el almacenamiento del carbono están influenciados por complejas interacciones de clima, suelo, manejo y especies vegetales.
Valores del coeficiente de correlación de Pearson (R) entre la biomasa y las variables de rendimiento para el bosque montano bajo, sur de Ecuador

*Valores de R superiores a 0,70 (positivo o negativo) indica que las variables están fuertemente correlacionadas, de 0,30 a 0,70 (positivo o negativo) indica que la correlación es moderada y de 0 a 0,30 se considera como débil correlación (RÜGNITZ et al., 2009).
Los modelos alométricos generados para estimar la biomasa aérea en el bosque montano bajo de la microcuenca El Padmi se indican en la Tabla 3, todos los modelos seleccionados indican un R2 ajustado superior al 80%, los parámetros de cada modelo fueron significativos (p<0,0001).
Modelos alométricos ajustados para el estrato arbóreo del bosque montano bajo del Sur de Ecuador
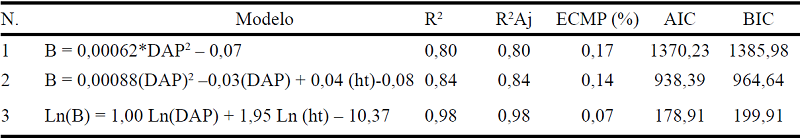
Donde: Nº = número de modelo alométrico generado; B= biomasa arbórea (t ha-1); N= 1408 individuos, DAP = diámetro a la altura del pecho (m); ht = altura total (m); G= área basal (m2); V= volumen (m3); R2= coeficiente de determinación, R2Aj = coeficiente de determinación ajustado; ECMP (%) = Error cuadrático medio ponderado; AIC= Criterio de información Akaike; BIC= Criterio de información Bayesiano.
En la Figura 2 se muestra la distribución gráfica del primer modelo generado y ajustado según se indica en la Tabla 3. Se realizó la representación gráfica del primer modelo generado debido a que tiene un buen coeficiente de determinación ajustado (R2Aj) y además utiliza una sola variable explicativa, el DAP, la cual es una de las variables dasométricas medidas en el campo con un alto valor de confiabilidad, como menciona Chave et al. (2005), el diámetro es la variable más importante en los modelos alométricos para la estimación de carbono. También, Nogueira et al. (2008) indican que utilizar el diámetro en modelos alométricos es una ventaja importante, para el uso práctico de la ecuación en la estimación de biomasa seca encima del suelo.
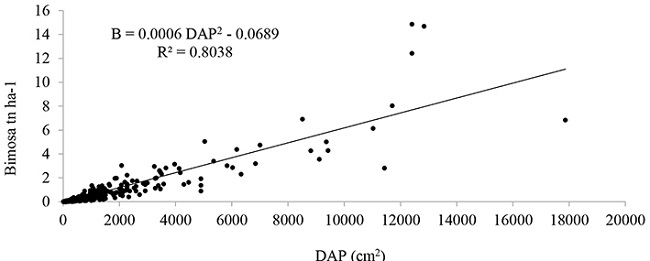
Figura 2
Relación entre la biomasa estimada y el diámetro a altura del pecho (DAP2), en arboles ≥ 5 cm de DAP del bosque montano bajo del Sur de Ecuador
Al comparar las tendencias de la acumulación de biomasa determinados en este estudio, con modelos generados para bosque montano bajo de otros lugares, como el modelo ajustado por Chave et al. (2005) y el modelo generado por Álvarez et al. (2012), se observó semejanzas en las tendencias de acumulación de biomasa en estos ecosistemas (Figura 3), esto indica que los modelos generados en esta investigación son factibles de aplicar como una herramienta para cuantificación de biomasa en ecosistemas tropicales. Además, según Montero y Montagnini (2005), el uso de ecuaciones alométricas permite el cálculo de la biomasa de una especie forestal de manera no destructiva y extrapolable a situaciones de crecimiento similares a las reales.
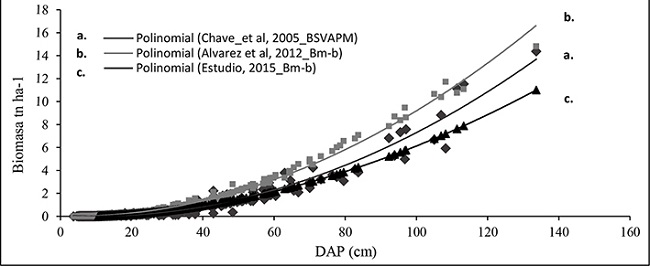
Figura 3
Representaciones de dispersión de la variable biomasa en bosques montanos bajos con: a) modelo alométrico generado por Chave et al. (2005); b) modelo alométrico generado por Álvarez et al. (2012); y c) modelo alométrico generado en este estudio
En el bosque montano bajo “El Padmi” existe diferencia estadística (p<0,05) entre dos grupos de familias botánicas según el almacenamiento de carbono en su biomasa, las variables que posiblemente expliquen esta división son: densidad de la madera, número de individuos por especies y el DAP, lo cual es corroborado por Fonseca, Alice y Rey (2009). En la Figura 4, se indica la media de almacenamiento de biomasa según los dos grupos taxonómicos diferenciados, el grupo uno en promedio almacena 0,05 t ha-1 y el grupo dos 0,28 t ha-1.
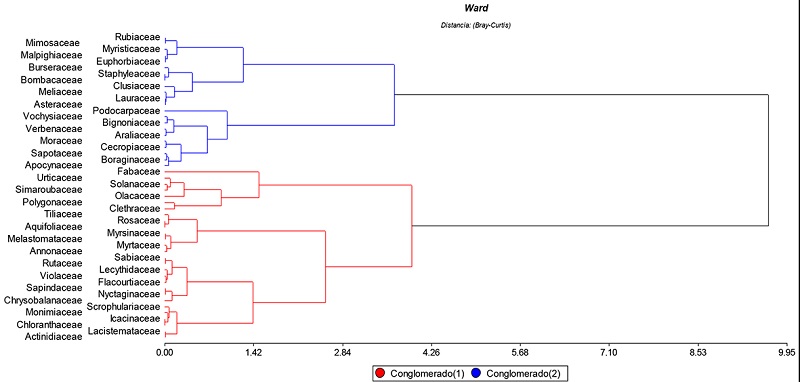
Figura 4
Dendrograma de las familias con mayor cantidad de carbono acumulado aplicando el método de Ward y distancia de Bray-Curtis
La familia que almacena la mayor cantidad de biomasa (74,83 t ha-1) y carbono (37,42 t C ha-1) es Moraceae. Las familias Moraceae, Lauraceae, Myristicaceae y Rubiaceae acumulan el 57% de carbono total (91,55 t C ha-1) del bosque montano bajo (Figura 5). Estos resultados son similares a los determinados en la amazonia peruana por Ureta (2015). Al referirnos a las especies botánicas, Pseudolmedia laevigata almacena la mayor cantidad de biomasa (55,23 t ha-1) y carbono (27,61 t C ha-1) de las 179 especies evaluadas. Esta especie, en conjunto con Nectandra reticulata, Otoba parvifolia, Vochysia paraensis, Ficus sp y Elaeagia obovata almacenan el 51% del carbono (81 t C ha-1) del bosque montano bajo (Figura 6). Los resultados encontrados demuestran que los bosques amazónicos almacenan elevadas cantidades de biomasa y carbono en su composición florística y estructura, sin embargo, esto depende de la fisiografía, tal y como lo atribuye Phillips et al. (1998) y Ureta (2015).
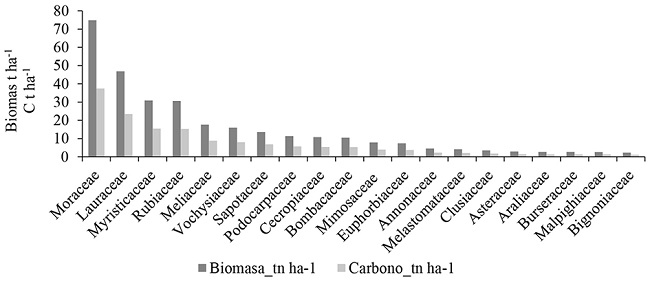
Figura 5
Familias con la mayor cantidad de carbono en el bosque montano bajo
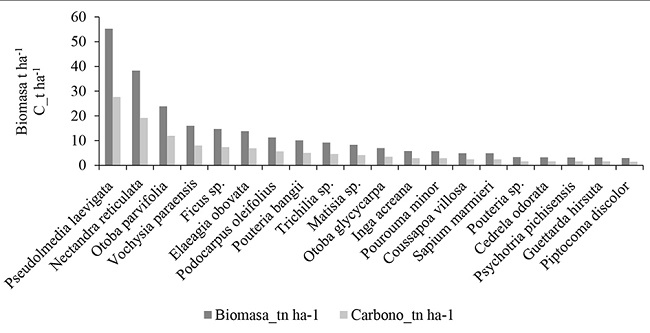
Figura 6
Especies botánicas con mayor cantidad de carbono en el bosque montano bajo
El bosque montano bajo almacena 161,07 t C ha-1 en 179 especies arbóreas y 52 familias.
Los modelos alométricos ajustados para predecir la biomasa en el estrato arbóreo del bosque montano bajo mostraron coeficientes de determinación (R ajustados) mayores al 80%. Las variables regresoras que mejor se ajustaron en la ecuación para la predicción de la biomasa aérea total fueron el diámetro a la altura de pecho al cuadrado (DAP2) y altura total (ht) tanto en escala lineal como logarítmica.
En el bosque montano bajo evaluado en el sur del Ecuador existen dos grupos de familias significativamente diferentes (p<0,05) de acuerdo al almacenamiento de carbono; siendo la familia Moraceae (74,83 t ha-1; 37,42 t C ha-1) y la especie Pseudolmedia laevigata Trécul (55,23 t ha-1; 27,61 t C ha-1) las que almacenan la mayor cantidad de biomasa y carbono respectivamente.
Los autores agradecen a la Dirección de Investigaciones de la Universidad Nacional de Loja por el financiamiento del proyecto de investigación “Evaluación del potencial de los servicios ecosistémicos de la microcuenca El Padmi, Zamora Chinchipe - Ecuador”, lo que permitió realizar este trabajo. A la comunidad “El Padmi” por la facilidad de los terrenos donde se instaló la parcela de estudio.
Figura 1
Ubicación geográfica del área de estudio, microcuenca “El Padmi, Zamora Chinchipe, sur de Ecuador
Biomasa total encima del suelo y reserva de carbono de la vegetación en el bosque montano bajo del sur de Ecuador
Valores del coeficiente de correlación de Pearson (R) entre la biomasa y las variables de rendimiento para el bosque montano bajo, sur de Ecuador
*Valores de R superiores a 0,70 (positivo o negativo) indica que las variables están fuertemente correlacionadas, de 0,30 a 0,70 (positivo o negativo) indica que la correlación es moderada y de 0 a 0,30 se considera como débil correlación (RÜGNITZ et al., 2009).
Modelos alométricos ajustados para el estrato arbóreo del bosque montano bajo del Sur de Ecuador
Donde: Nº = número de modelo alométrico generado; B= biomasa arbórea (t ha-1); N= 1408 individuos, DAP = diámetro a la altura del pecho (m); ht = altura total (m); G= área basal (m2); V= volumen (m3); R2= coeficiente de determinación, R2Aj = coeficiente de determinación ajustado; ECMP (%) = Error cuadrático medio ponderado; AIC= Criterio de información Akaike; BIC= Criterio de información Bayesiano.
Figura 2
Relación entre la biomasa estimada y el diámetro a altura del pecho (DAP2), en arboles ≥ 5 cm de DAP del bosque montano bajo del Sur de Ecuador
Figura 3
Representaciones de dispersión de la variable biomasa en bosques montanos bajos con: a) modelo alométrico generado por Chave et al. (2005); b) modelo alométrico generado por Álvarez et al. (2012); y c) modelo alométrico generado en este estudio
Figura 4
Dendrograma de las familias con mayor cantidad de carbono acumulado aplicando el método de Ward y distancia de Bray-Curtis
Figura 5
Familias con la mayor cantidad de carbono en el bosque montano bajo
Figura 6
Especies botánicas con mayor cantidad de carbono en el bosque montano bajo
