Research articles
Updated list and new records of macroalgae from the Gulf of Morrosquillo, Colombian Caribbean
Listado actualizado y nuevos registros de macroalgas para el Golfo de Morrosquillo, Caribe colombiano
 Diego Alexander Hernández-Contreras
Brigitte Gavio
Diego Alexander Hernández-Contreras
Brigitte Gavio
Updated list and new records of macroalgae from the Gulf of Morrosquillo, Colombian Caribbean
Acta botánica mexicana, no. 131, e2288, 2024
Instituto de Ecología A.C., Centro Regional del Bajío
Received: 23 November 2023
Revised document received: 08 January 2024
Accepted: 01 March 2024
Published: 19 April 2024
Abstract
Background and Aims: Macroalgae are primary producers of marine ecosystems, so the information regarding their species composition and distribution is necessary to develop conservation strategies and sustainable use of the coastal coastlines they inhabit. However, in different marine regions of Colombia, such as the Gulf of Morrosquillo, this knowledge is scarce, requiring a greater sampling effort and constant updating of data. Therefore, this work aims to provide an updated list of the macroalgae in the aforementioned region, based on published and unpublished biological records and recent sampling.
Methods: Bibliographic information was compiled from historical records from 1976 to the present and complemented with sampling in one location in the Gulf of Morrosquillo. The taxa were organized and updated based on the most recent nomenclature, and the geographical affinity of the algal flora was determined using the Cheney index.
Key results: The occurrence of 97 taxa classified in three classes, 15 orders, 32 families and 54 genera. Red algae, with 48 taxa, were the group that contributed the largest number of species, and the phycoflora of the region was of a mixed type according to the Cheney index. Six new annotated and illustrated records were reported for the Gulf: Antithamnionella breviramosa, Centroceras gasparrinii, Bostrychia tenella, Melanothamnus sphaerocarpus, Agardhiella ramosissima and Feldmannia mitchelliae.
Conclusions: The inventory of macroalgae in the evaluated region was updated, showing a clearer historical panorama of the sampled areas, which suggests an extraordinary potential in the future discovery of new records that would increase the knowledge about the phycoflora of the Colombian Caribbean, particularly because the records of encrusting red algae, epiphytic and epizoic species are non-existent.
Key words: Biodiversity+ Chlorophyta+ Heterokontophyta+ review+ Rhodophyta+ taxonomy.
Resumen
Antecedentes y Objetivos: Las macroalgas son productores primarios de los ecosistemas marinos, por lo que la composición de especies y su distribución es información necesaria para establecer estrategias de conservación y uso sostenible de los litorales costeros que habitan. Sin embargo, en diferentes regiones marinas de Colombia, como el Golfo de Morrosquillo, este conocimiento es escaso, requiriendo de un mayor esfuerzo de muestreo y actualización constante de datos. Por lo tanto, este trabajo tiene como objetivo proporcionar una lista actualizada de las macroalgas de la región mencionada, basada en registros biológicos publicados, inéditos y muestreos recientes.
Métodos: Se recopiló información bibliográfica de registros históricos desde 1976 al presente y se complementó con muestreos en una localidad del Golfo de Morrosquillo. Los taxa se organizaron y actualizaron con base en la nomenclatura más reciente, y se determinó la afinidad geográfica de la flora algal mediante el índice de Cheney.
Resultados clave: Se determinó la presencia de 97 taxa agrupados en tres clases, 15 órdenes, 32 familias y 54 géneros. Las algas rojas, con 48 taxa, fueron el grupo que aportó un mayor número de especies, y la ficoflora de la región es de tipo mixto de acuerdo con el índice de Cheney. Se reportan seis nuevos registros comentados e ilustrados para el Golfo: Antithamnionella breviramosa, Centroceras gasparrinii, Bostrychia tenella, Melanothamnus sphaerocarpus, Agardhiella ramosissima y Feldmannia mitchelliae.
Conclusiones: Se actualizó el inventario de macroalgas en la región evaluada, mostrando un panorama histórico más claro de las zonas muestreadas, lo que sugiere un extraordinario potencial en el hallazgo futuro de nuevos registros que incrementarían el conocimiento sobre la ficoflora del Caribe colombiano, particularmente porque los registros de algas rojas incrustantes, especies epífitas y epizoicas son nulos.
Palabras clave: Biodiversidad, Chlorophyta, Heterokontophyta, revisión, Rhodophyta, taxonomía.
Introduction
Seaweeds play a prominent role in coastal marine ecosystems as primary producers (Peña-Salamanca, 2008; Tuya et al., 2014); even so, despite their ecological importance, many regions in the world still have a limited knowledge of the biodiversity, physiology, and taxonomy of macroalgae along their coasts (Hu and Fraser, 2016). These benthic organisms fix carbon through photosynthetic processes, producing organic matter which is exported to other environments and contribute to the nutrient cycle in marine systems (Goreau, 1963; Bykova et al., 2020), and to blue carbon sequestration (Ortega et al., 2019; Queirós et al., 2019). Macroalgae are used as a refuge by larval stages of commercial fish and invertebrates (Vandendriessche et al., 2007; Tano et al., 2016). Several seaweed taxa are an important economic resource of phycocolloids and other active compounds used in food, cosmetic and pharmaceutical industries (Zerrifi et al., 2018; Hurtado et al., 2019). These characteristics, together with their ability to structure complex biological communities, convert them in bioengineer organisms in intertidal ecosystems (Umanzor et al., 2019).
In the history of Marine Ecology and Phycology of Colombia, one of the first floristic lists of seaweeds in the Colombian Caribbean was made by Germán A. Bula Meyer, who listed 472 species inhabiting this ecoregion of the country (Bula-Meyer, 1998). However, he did not record macroalgae from the Gulf of Morrosquillo. Subsequently, Díaz-Pulido and Díaz-Ruíz (2003) reported 29 species of macroalgae for the Gulf, and this species list has not been updated since then.
The studies of the marine algal flora of the Caribbean coast of Colombia have been focused on the eastern area towards Santa Marta (Schnetter, 1969; Bula-Meyer and Díaz-Pulido, 1995; Díaz-Pulido and Díaz-Ruíz, 2003; Vega-Sequeda et al., 2008; Camacho and Montaña-Fernández, 2012; Gómez-Cubillos, 2018; Cabarcas and García-Ureña, 2020), Rosario islands near Cartagena (Salazar-Forero et al., 2021), and La Guajira peninsula (Díaz-Pulido, 2000; Mendoza-Castro and Suarez-Acuña, 2018; Vasquez-Carrillo and Sullivan, 2018; Puccini and Molina, 2022; Gavio et al., 2022b). Phycological research in the International Biosphere Reserve Seaflower, oceanic archipelago located in the western Caribbean, has been extensive (e.g., Díaz-Pulido and Bula-Meyer, 1997; Ortiz and Gavio, 2010; Albis-Salas and Gavio 2011, 2015; Rincón-Díaz, 2014; Gavio et al., 2015; Rincón-Díaz and Ramos, 2016; Rincón-Díaz et al., 2018; Reyes-Gómez et al., 2021; Gavio et al., 2022a; Daza, 2023).
In contrast, the western part of the Caribbean coast of the country (with exception of work done by López-Jiménez et al. (2021) in the Capurganá Bay, who reported 61 species of macroalgae inhabiting this region) and especially the Gulf of Morrosquillo, located between the departments of Córdoba and Sucre (Castaño, 2002), has been overlooked and is lacking adequate knowledge of its phycoflora (Gavio, 2023), as shown by the low number of reported species by Díaz-Pulido and Díaz-Ruíz (2003). However, some authors have reported macroalgal species for both rocky (Quirós-Rodríguez et al., 2010, 2013; Sampedro-M. et al., 2012) and sandy shores (Quintana-M. and Mercado-Gómez, 2017) of the Gulf following the list of 2003. In Colombia, this lack of knowledge of the phycoflora of its coasts is due to several factors: first of all, despite having coasts on both the Atlantic and the Pacific Ocean, the country does not value its marine ecosystems as it should (Garnica, 2021); second, historically the study of marine algae does not rank high in the interest of the national scientific community, although, at present, this tendency is changing (Gavio, 2023); third, the access to several areas of coastline is still difficult and expensive, either due to lack of infrastructure (roads and highways), or security problems due to the presence of paramilitary groups or other illegal associations in some territories (Garnica, 2021; Palencia et al., 2023).
In accordance with the ecological importance of seaweeds and the lack of updated information on the phycoflora that inhabits the Gulf of Morrosquillo, the purpose of this paper was to provide an updated list of the macroalgae of the Gulf, through the review of existing and unpublished literature, along with recent samplings. It is intended that this information contributes to better decision-making in marine-coastal management of the Colombian Caribbean.
Materials and Methods
Macroalgal samples were collected in January, April, and September 2019 and January 2020 around the town of Coveñas (department of Sucre) in the Gulf of Morrosquillo (Fig. 1, the square on the last map indicates the study zone). The Gulf lies in a warm dry climate, with a mean annual temperature of 27.4 °C and annual precipitation ranging between 82.9 and 131 mm (Aguilera-Díaz, 2005). The Gulf harbors sandy substrate and coral patches, while the shore is characterized by the presence of mangrove and coastal lagoons (Díaz-Merlano and Puyana, 1994).
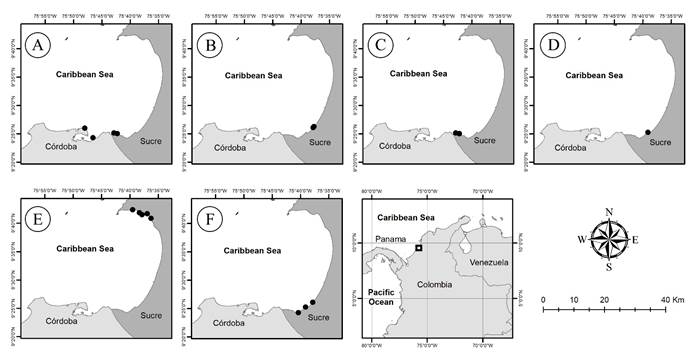
Figure 1:
Sampling sites in different studies in the Gulf of Morrosquillo, Colombia. A. Quirós-Rodríguez et al. (2010); B. Sampedro-M. et al. (2012); C. Quirós-Rodríguez et al. (2013); D. Meneses and Rivera (2014); E. Quintana-M. and Mercado-Gómez (2017); F. new records for this study. The works by Schnetter (1976; 1978), Patiño and Flórez (1993), and Díaz-Pulido and Díaz-Ruíz (2003) were not added to the map because they do not have the coordinates, making their location in the Gulf uncertain.
Macroalgae were sampled in inter- and subtidal environments to a depth of about 2 m. Specimens were preserved in a formalin-seawater solution at 5% (Gavio et al., 2015) until their identification, and were deposited with inclusion codes (IC), or vouchers, in the Pacific Oceanographic and Hydrographic Research Center biological collection (CB-CCCP), Tumaco, Colombia, Single National Collection Registry under number 138 of the Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. For some specimens, transverse and longitudinal sections were made by hand with a razor blade and stained with aniline blue at 1% (Florez-Leiva et al., 2010). The specimens were identified with specific literature (e.g., Taylor, 1960; Littler and Littler, 2000; Dawes and Mathieson, 2008; Littler et al., 2008; Won et al., 2009) under an optical microscope (Labomed LX 400, Fremont, USA). The photographs were taken in the biology laboratory of the Pacific Oceanographic and Hydrographic Research Center, and in the Imaging Laboratory of the Postgraduate program in Sciences-Biology, Universidad del Valle (Cali, Colombia).
Records from published papers were included to prepare the species list from Schnetter (1976) to the most recent work in the Gulf by Quintana-M. and Mercado-Gómez (2017), and books and theses available in the libraries of the following national institutions were consulted: Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis (INVEMAR), Universidad de Córdoba, Universidad Jorge Tadeo Lozano, Universidad del Magdalena and Universidad de Sucre. Additionally, the online site macroalgae.org (Macroalgal Herbarium Portal, 2024) and the data available in the Global Biodiversity Information Facility (GBIF) from the virtual Herbarium of Universidad Nacional de Colombia (COL) (Raz and Agudelo-Zamora, 2023) were consulted. The scientific names were corroborated in AlgaeBase (Guiry and Guiry, 2024) and ITIS (2024). The taxonomic categories in the checklist were organized according to Wynne (2022). Nevertheless, the list does not include the name Ochrophyta, given the recent validation of the phylum Heterokontophyta (Guiry et al., 2023).
The information was filtered with the tool “taxon match” in the world record of marine species (WoRMS, 2023) and the software OpenRefine v. 3.4.1 (Verborgh and De Wilde, 2013). Taxonomic notes were provided, when necessary, regarding the nomenclatural updates or the new records. The geographical map was elaborated with the software QGIS v. 3.34 (QGIS, 2023) (Fig. 1). In order to determine the geographical affinity of the phycoflora, the Cheney index was calculated:
where R is the number of red algal taxa, C is the number of green algae, and P is the number of brown algae. Values <3 indicate cold temperate flora, while values >6 indicate tropical flora. Intermediate values indicate mixed flora (e.g. warm temperate flora) (Cheney, 1977).
Results
Phycoflora diversity
A total of nine publications with macroalgal reports for the Gulf of Morrosquillo were retrieved (Table 1, Fig. 1A-E): five published articles, one thesis and three books. Among these works, Schnetter (1976; 1978), Díaz-Pulido and Díaz-Ruíz (2003), and Quirós-Rodríguez et al. (2010) were the only ones that have the objective of presenting a floristic list for the Caribbean coast of Colombia or the coast of Córdoba, including the Gulf of Morrosquillo. The other works, namely Patiño and Flórez (1993), Sampedro-M. et al. (2012), Quirós-Rodríguez et al. (2013), Meneses and Rivera (2014), and Quintana-M. and Mercado-Gómez (2017)) had other study objectives (ecological), and macroalgal species records were an additional result.
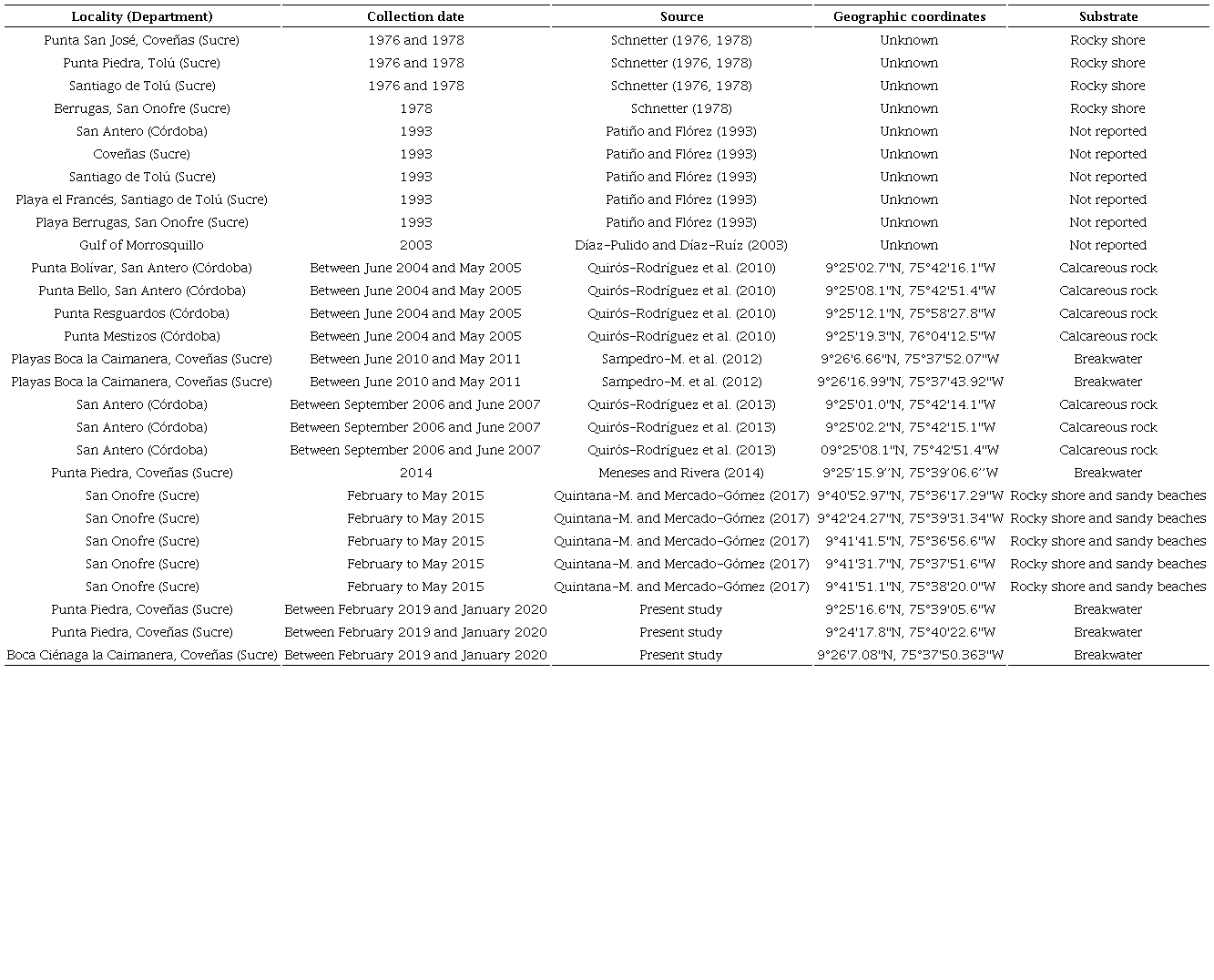
A total of 97 taxa are reported, corresponding to 96 species and one infraspecific name (Appendix). Species are distributed in three classes, 15 orders, 32 families and 54 genera (Fig. 2). The brown algae (Heterokontophyta) present the lowest number of species (16), of which twelve are members of the family Dictyotaceae. Red algae (Rhodophyta) are composed of 48 taxa. The family Gracilariaceae is the most representative within the red algae with 11 species. Finally, the green algae (Chlorophyta) contribute with 33 taxa. The most diverse family is the Caulerpaceae with eight taxa (seven species and one form). Twenty-one years later, the present work adds 68 records not included in the species list by Díaz-Pulido and Díaz-Ruíz (2003).
Figure 2:
Number of records in the Gulf of Morrosquillo, Colombia, according to different taxonomic levels.
New records for the Gulf of Morrosquillo
From the species presented, six correspond to new records for the Gulf of Morrosquillo (Fig. 1f), five red algae: Antithamnionella breviramosa (E.Y. Dawson) Wollaston, Centroceras gasparrinii (Meneghini) Kützing, Bostrychia tenella (J.V. Lamouroux) J. Agardh, Melanothamnus sphaerocarpus (Børgesen) Díaz-Tapia & Maggs, Agardhiella ramosissima (Harvey) Kylin, and the brown alga Feldmannia mitchelliae (Harvey) H.-S. Kim. Their description and illustrations are presented hereafter.
Taxonomy
HETEROKONTOPHYTA
Phaeophyceae
Ectocarpales
Acinetosporaceae
Feldmannia mitchelliae (Harvey) H.-S. Kim, Algal flora of Korea 2(1): 3-137. 2010. Figs. 3A-C.
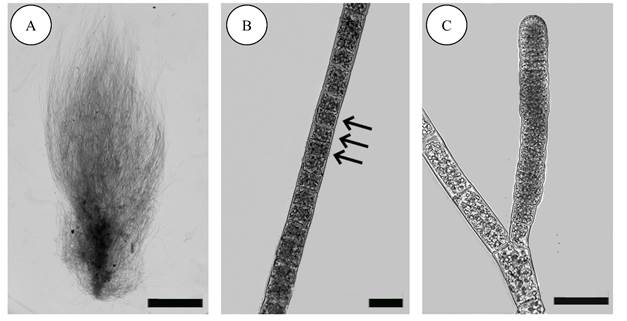
Figure 3:
A-C. Feldmannia mitchelliae (Harvey) H.-S. Kim. A. habit; B. intercalary meristem (arrows); C. plurilocular gametangium. Scale bars: A=2 mm, B=20 µm, C=50 µm.
TYPE: UNITED STATES OF AMERICA. Massachusetts, Nantucket, no depth and coordinate information available, 1987, H. B. S. Womersley 52 (holotype: TCD).
Thallus filamentous, brown, 8 cm tall, entirely uniseriate, branching irregular, axial cells 15-25 µm diameter, 30-80 µm long, branches tapering towards apex, roundish (Fig. 3A); cell divisions in intercalary meristem (Fig. 3B); plurilocular gametangia cylindrical, 15-30 µm diameter, 70-130 µm long, sessile, inserted laterally on branches (Fig. 3C).
Taxonomic notes: it can be differentiated from other species of the genus by not presenting branches at an 90° angle, the filaments gradually decrease in size until reaching the apex and its sporangia are mainly sessile (not stalked) and plurilocular.
Distribution in Colombian Caribbean: Guajira, Tayrona National Natural Park (Tayrona-NNP), Magdalena, Darién, Archipelago of San Andres, Old Providence and Sainte Cataline and Gulf of Morrosquillo.
Material examined: COLOMBIA. Sucre, Coveñas, Boca Ciénaga la Caimanera, depth 1 m, 9°26'7.08''N, 75°37'50.363''W, 10.I.2020, H. L. Quintana-M. Macroalg-CCCP-07 (CB-CCCP).
RHODOPHYTA
Florideophyceae
Ceramiales
Ceramiaceae
Antithamnionella breviramosa (E.Y. Dawson) Wollaston, Philosophical Transactions of the Royal Society of London, B. Biological Sciences 259: 257-352. 1970. Fig. 4A.

Figure 4:
A. Antithamnionella breviramosa (E.Y. Dawson) Wollaston. A. close up of erect axis with gland cells (arrows) touching only one branch cell; B-C. Centroceras gasparrinii (Meneghini) Kützing. B. forcipate apices; C. close-up of nodes with spines; D-I. Bostrychia tenella (J.V.Lamouroux) J.Agardh. D. habit; E. branch, showing monosiphonic branchlets (arrow); F. cortication of axes; G. transverse section of thallus; H. urn-shaped cystocarp; I. branch with stichidia (arrow). Scale bars: A=20 µm, B, C=50 µm, D=2 mm, E=0.25 mm, F=150 µm, G=100 µm, H=300 µm, I=100 µm.
TYPE: UNITED STATES OF AMERICA. California, Santa Catalina Island, Pebbly Beach, near Avalon, no depth and coordinate information available, 24.III.1948, E. Y. Dawson 26521 (holotype: AHFH).
Thallus filamentous, delicate, 1.5 mm tall, rose-pink in color; prostrate axes uniseriate, 20-30 µm diameter, bearing rhizoids; erect axes uniseriate, 15-30 µm diameter, 5-22 cells long, cells 25-35 µm long; branchlets in whorls of three, at the upper end of axial cells, tapering from base toward apex, each branchlet 10-12 µm diameter, divided pseudodichotomously; gland cells oval, 10-12 µm diameter, in contact with only one cell of branchlets (Fig. 4A).
Taxonomic notes: Kim and Lee (1990) determined that the number of whorls branchlets arising from each axial cell is a good character to identify this species. For Antithamnionella breviramosa, they observed that the alga always forms three branchlets per axial cell; this character was observed in our specimen.
Distribution in Colombian Caribbean: Guajira, Tayrona-NNP, Darién, Archipelago de San Andrés, Old Providence and Sainte Cataline and Gulf of Morrosquillo.
Material examined: COLOMBIA. Sucre, Coveñas, Punta Piedra, depth 2.5 m, 9°25'16.6''N, 75°39'05.6''W, 5.I.2019, epiphytic on Halimeda opuntia (L.) J.V. Lamouroux, H. L. Quintana-M. Macroalg-CCCP-02 (CB-CCCP).
Centroceras gasparrinii (Meneghini) Kützing, Lipsiae: F.A. Brockhaus (i)-vi, 1-922. 1849. Figs. 4B, C.
TYPE: ITALY. Sicilia, Palermo, no depth and coordinate information available, 1844, G. Meneghini 186 (holotype: L-0193946, isotype: L-0193946, #940264-102).
Thallus filamentous, red to brown in color, 5 cm tall; axial filament completely corticated by small rectangular cells in vertical row; branching dichotomous or occasionally trichotomous, apices weakly incurved (Fig. 4B); branches 100-120 µm diameter; nodes bearing spines formed by two or three cells, acropetal cells ovoid: gland cell spherical at nodes (Fig. 4C); nodes 120-150 µm diameter, 12-15 pericentral cells.
Taxonomic notes: distinguishable among other species of this genus by the presence of ovoid terminal acropetal cells.
Distribution in Colombian Caribbean: Tayrona-NNP, Magdalena and Gulf of Morrosquillo.
Material examined: COLOMBIA. Sucre, Coveñas, Boca Ciénaga la Caimanera, depth 1 m, 9°26'7.08''N, 75°37'50.363''W, 05.I.2019, H. L. Quintana-M. Macroalg-CCCP-03 (CB-CCCP).
Rhodomelaceae
Bostrychia tenella (J.V. Lamouroux) J. Agardh, Lundae (Lund): C. W. K. Gleerup 2(3): 787-1138, 1158-1291. 1863. Figs. 4D-I.
TYPE: UNITED STATES OF AMERICA. Virgin Islands, Saint Croix, no depth and coordinate information available, 1998, T. Yoshida 672309A (syntype: MEL).
Thalli forming mats up to 2 cm tall, purple, or brown in color, growing on mangrove roots (Fig. 4D); abundant branching, pinnate branches emerging in alternating fashion, branchlet apices upturned, branches 170-300 µm diameter, without cortication, superficial cells small, pit connection present, branchlets 80-120 µm diameter, branches uniseriate at the apex, 20-50 µm diameter (Fig. 4EE; main axes with light cortication (Fig. 4F); single central cell surrounded by 5-9 pericentral cells in main axes, two tier cells per pericentral cell (Fig. 4G); cystocarps urn-shaped, sessile, 600-650 µm diameter (Fig. 4H); stichidia 500-600 µm high, 100-150 µm diameter (Fig. 4I).
Taxonomic notes: this species forms a cryptic complex with the taxon Bostrychia binderi Harvey (Zuccarello et al., 2015), and its morphological differentiation is rather difficult (Saengkaew et al., 2016; Zuccarello et al., 2015). However, authors such as Hernández-Contreras (2017) suggested that these two species could be distinguished by the fact that B. tenella has completely monosiphonous branchlets, with upturned apices and light cortication.
Distribution in Colombian Caribbean: Tayrona-NNP, Islas del Rosario National Natural Park, Darién, and Gulf of Morrosquillo.
Material examined: COLOMBIA. Sucre, Coveñas, Boca Ciénaga la Caimanera, depth 0 m, 9°26'7.08''N, 75°37'50.363''W, 05.I.2019, epiphytic on Rhizophora mangle L. roots H. L. Quintana-M. Macroalg-CCCP-04 (CB-CCCP).
Melanothamnus sphaerocarpus (Børgesen) Díaz-Tapia & Maggs, European Journal of Phycology 52(1): 1-20. 2017. Figs. 5A-E.
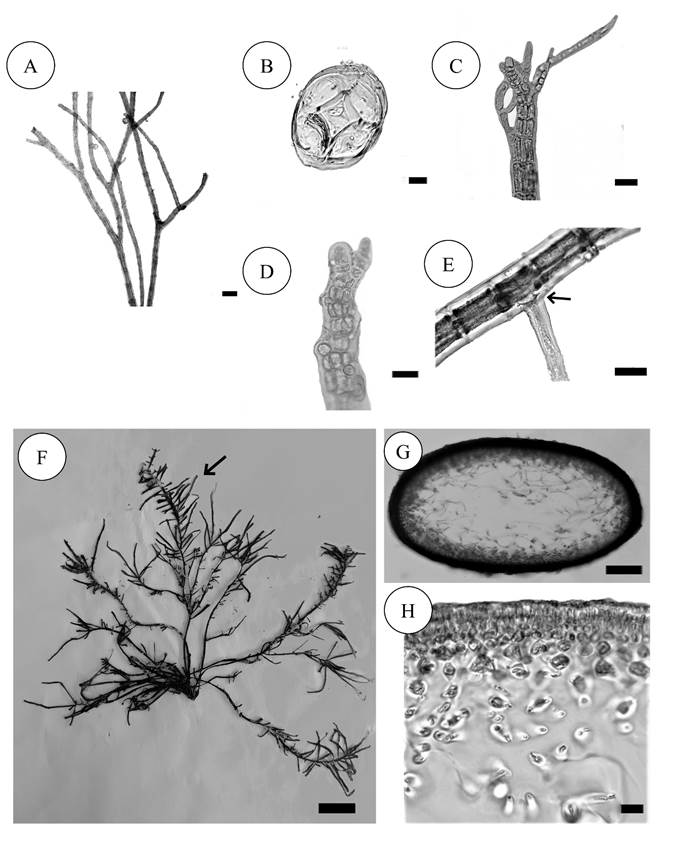
Figure 5:
A-E. Melanothamnus sphaerocarpus (Børgesen) Díaz-Tapia & Maggs. A. habit, showing dichotomous and pseudodichotomous branching; B. transverse section of thallus showing four pericentral cells; C. apical portion with trichoblast; D. apical portion showing primordial arrangement; E. prostrate axis with rhizoid cut off from pericentral cells (arrow); F-H. Agardhiella ramosissima (Harvey) Kylin. F. habit, showing opposite branching (arrow); G. transverse section of thallus showing compressed axis, with transverse filament in the center; H. close up transverse section of thallus showing cortical cells. Scale bars: A=100 µm, B=10 µm, C=50 µm, D=10 µm, E= 50 µm, F=2 cm, G=300 µm, H=10 µm.
TYPE: UNITED STATES OF AMERICA. Virgin Islands, Store Nordsidebugt (Magens Bay), St. Thomas, no depth and coordinate information available, 1918, F. Børgesen (holotype: C?, isotype: US, C).
Thallus filamentous, dark red, forming mats up to 1 cm tall on the substrate, prostrate (60-85 µm diameter, segments 1-2 diameter long) and erect axes, dichotomously branched, 40-50 µm diameter (Fig. 5A); axes not corticated, with four pericentral cells (Fig. 5B); trichoblasts at the apices (Fig. 5C); apex with monosiphonic cells (Fig. 5D); rhizoids unicellular, cut off from pericentral cell (Fig. 5E).
Taxonomic notes: this species is characterized by its pseudodichotomous branching and thallus with a small size of no more than 2 cm high.
Distribution in Colombian Caribbean: Darién, Gulf of Morrosquillo, Archipelago of San Andres, Old Providence and Sainte Cataline.
Material examined: COLOMBIA. Sucre, Coveñas, Boca Ciénaga la Caimanera, depth 0 m, 9°26'7.08''N, 75°37'50.363''W, 05.I.2019, growing on limestone rock H. L. Quintana-M. Macroalg-CCCP-05 (CB-CCCP).
Gigartinales
Solieriaceae
Agardhiella ramosissima (Harvey) Kylin, Acta Universitatis Lundensis 28(8): 1-88. 1932. Figs. 5F-H.
TYPE: UNITED STATES OF AMERICA. Florida, Key West, no depth and coordinate information available, 1853, W.H. Harvey (holotype: TCD?).
Thallus cartilaginous, 20 cm tall, pink, green, main axes flat, 2-3 mm wide, 1 mm thick, branching irregular proximally and pinnate distally, branchlets tapering toward the apices and at the base (Fig. 5F); medulla formed by dispersed filaments 1-2 µm diameter in a mucilaginous matrix, cortex formed by three to four layers of deeply pigmented cells (Figs. 5G, H).
Taxonomic notes: this taxon is easily distinguishable from Agardhiella subulata (C. Agardh) Kraft & M.J. Wynne due to its flattened thallus and the pinnate branching.
Distribution in Colombian Caribbean: Darién and Gulf of Morrosquillo.
Material examined: COLOMBIA. Sucre, Coveñas, Punta piedra, depth 1 m, 9°24'17.8''N, 75°40'22.6''W, 05.I.2019, growing on limestone rock H. L. Quintana-M. Macroalg-CCCP-06 (CB-CCCP).
Climate affinity of the marine flora of Gulf of Morrosquillo
The Cheney index was calculated with the updated list of species for the Gulf of Morrosquillo. The value of the index was 5.02, indicating a mixed flora, in transition between temperate and tropical affinities.
Discussion
Richness of species
This work contributes to the marine-coastal management of the Gulf of Morrosquillo with an updated taxonomic list that records the presence of at least 97 taxa of seaweeds for the region, since Díaz-Pulido and Díaz-Ruíz (2003), in the first checklist of the Gulf, previously reported the existence of 29 species (this list and others that precede it include some records in personal collections, which was common practice at that time, since the legalization of biological collections in Colombia was an incipient policy). This represents an increase of 235% in the reported species between the two works, which indicates that during 21 years the generation of new phycological knowledge for the region, despite logistical and security difficulties, has been expanded. To date, seaweed sampling in the Gulf shows a greater effort in its southern portion (Fig. 1, Table 1), around the municipalities of San Antero and Coveñas. This might be associated with better access facilities, vicinity to towns and lack of guerrilla activity in these areas. On the contrary, the middle and northern parts of the Gulf lack access roads, towns on the coast, and there is the presence of paramilitary groups still controlling the territory, all of which complicate sampling.
The majority of the works reported in the present paper were not focused on macroalgae, reporting algal species as additional results, reinforcing the evidence that algal species richness in the Gulf is underestimated. Furthermore, the lack of records of incrusting red algae, epiphytic, epizoic species and cyanobacteria is noteworthy. These organisms constitute a relevant portion of macroalgal diversity, especially in tropical regions (Mateo-Cid et al., 2020; Reyes-Gómez et al., 2021) and have been ignored in the Gulf of Morrosquillo as well as in the Colombian Caribbean in general. Moreover, most recent works (e.g., Quirós-Rodríguez et al., 2010; Sampedro-M. et al., 2012; Meneses and Rivera, 2014; Quintana-M. and Mercado-Gómez, 2017) focused exclusively on shallow environments (to 4 m depth); hence, there is no information on seaweeds at greater depths in the Gulf.
New records of seaweeds
The new records of this work and other comments on some species of interest for the coastal management of the Gulf of Morrosquillo considered the current nomenclature and taxonomic changes reported by different authors. Until 2009, the most common species of the genus Centroceras Kützing reported for the Caribbean was C. clavulatum (C. Agardh) Montagne. However, Won et al. (2009) showed that the distribution of this species is restricted to the Southern Pacific Ocean. To date, at least nine different species are present in the Western Atlantic Ocean, according to Wynne's most recent review (2022): C. arcii C.W. Schneider, Cianciola & Popolizio, C. gasparrinii (Meneghini) Kützing, C. hyalacanthum Kützing, C. illaqueans C.W. Schneider, Cianciola & Popolizio, C. internitens S.G. Gallagher & Humm, C. micracanthum Kützing, C. minutum Yamada, C. rhizophorum Montagne, C. tetrachotomum Won, T.O. Cho & Fredericq (Schneider et al., 2015; Wynne, 2022). Considering these studies, we propose discounting reports of the species C. clavulatum from the species list of the Gulf of Morrosquillo (and the country). Recent samplings in other regions of the Colombian coast have shown that C. gasparrinii is the most common species of the genus (Gavio, unpublished results).
Other records for the Gulf, such as Bostrychia tenella, stand out due to the complexity of their identification. Given its great phenotypic plasticity, Zuccarello and West (2011) group the species into complexes, the Bostrychia tenella / B. binderi complex being one of them. Zuccarello et al. (2015) and Hernández-Contreras (2017) have elucidated some characters that are useful to differentiate these taxa using molecular tools and comparing the morphology of the species.
The sample found in the Gulf of Morrosquillo (H. L. Quintana-M. Macroalg-CCCP-04) belongs to the mentioned complex, due to the presence of two characters that helped to identify this species. First, the presence of numerous monosiphonous branches; this character has been one of the most prominent to differentiate the species of this complex since B. binderi presents few or no monosiphonous branches, while B. tenella presents a large number of these branches arranged in a revolute fashion giving it a feather-like appearance (Fig. 4E shows the presence of these structures being similar to images 7, 8 and 9 of Zuccarello et al. (2015) and figure B of image 5 in Saengkaew et al. (2016). Second, the level of cortication of the thallus: B. tenella has moderate cortication in the main axes (Fig. 4F), while in lateral branches and immature axes there is no presence of cortical cells. On the contrary, B. binderi presents dense cortication both in main axes and in branches and branchlets, see Figs. 1-6 in Zuccarello et al. (2015), figures F and G in Saengkaew et al. (2016) and figure I of Hernández-Contreras (2017).
Hernández-Contreras (2017) highlighted the absence of cladohaptera in B. tenella as an important feature to differentiate this species from B. binderi, in which cladohaptera usually are present. Nevertheless, Zuccarello et al. (2015), Saengkaew et al. (2016), and Hernández-Contreras (2017) did not always report the absence or presence of cladohaptera in their specimens (which depends on the sampling conditions), which makes this character less informative than expected for the differentiation between the two species, compared to the number of monosiphonic branches, and especially, the cortication of the thallus. This is so, not only because of the typical crypticism observed in the genus Bostrychia Montagne (Zuccarello and West, 2011), but also because B. tenella is a species with high genetic diversity, and complex phylogeographic patterns, which have been explained by the effect of geological history and marine surface currents (Zuccarello et al., 2015; Bulan et al., 2022), which can eventually make it very susceptible to generating morphological variations that complicate its identification by classical taxonomical studies.
Regarding the other new records for the Gulf of Morrosquillo, apart from some sporadic and discontinuous records for Agardhiella ramosissima in the Darién (Díaz-Pulido and Díaz-Ruíz, 2003), and for Feldmannia mitchelliae in the north of La Guajira peninsula (Díaz-Pulido and Díaz-Ruíz, 2003; Vasquez-Carrillo and Sullivan, 2018), little is known about the biology or ecology of these species in the Colombian Caribbean context. Nevertheless, Antithamnionella breviramosa has a relatively greater number of records concentrated in the International Biosphere Reserve Seaflower (Díaz-Pulido and Bula-Meyer, 1997; Rincón-Díaz, 2014; Rincón-Díaz and Ramos, 2016; Gavio and Daza, 2022), and some occasional reports in Darién (Díaz-Pulido and Díaz-Ruíz, 2003), Santa Marta (Bula-Meyer and Díaz-Pulido, 1995; Díaz-Pulido and Díaz-Ruíz, 2003), and La Guajira Peninsula (Díaz-Pulido, 2000). Melanothamnus sphaerocarpus has only been reported on the island of San Andrés by Ortiz and Gavio (2010) in the drift, and Albis-Salas and Gavio (2015) in Thalassia grasslands, who described it based on a specimen in a reproductive state and named it Neosiphonia sphaerocarpa (Børgesen) M.-S. Kim & I.K. Lee, a name that is currently considered synonymous of M. sphaerocarpus (Wynne, 2022; Guiry and Guiry, 2024).
Climate affinity
It is well-known that in the tropics the marine phycoflora is dominated by red algae, followed by green algae (Hu and Fraser, 2016). Brown seaweeds have a reduced diversity at low latitudes, while they are dominant in cold and temperate waters (Mathieson et al., 2009; Neiva et al., 2016). This knowledge is captured in the Cheney index, a simple way to calculate biogeographic affinities of a local/regional marine flora. According to Cheney (1977) and Mathieson et al. (2009), this index is described as: “(R+C)/P, where R=the number of Rhodophyta, C=the number of Chlorophyta, and P=the number of Phaeophyceae, (or Heterokontophyta). A value of <3.0 indicates a temperate or cold-water flora, while values of >6.0 indicate a tropical flora”. Intermediate values are associated with flora of a mixed type that generally inhabits warm temperatures (Mathieson et al., 2009). The result of this ratio for the flora of Gulf of Morrosquillo is a mixed flora, despite the high temperatures of the Gulf waters (28-29.5 °C) and its location in the tropics (Bastidas-Salamanca and Ordóñez-Zúñiga, 2017). This indicates a low representation of red algae, which are mainly small, epiphytic, and therefore are easily overlooked. Since the majority of the recent literature was not especially focused on algae, the absence of these records is justified. The lack of specific and long-term studies is the most probable reason for the low representation of red algae.
Research perspectives in the Gulf of Morrosquillo
Caulerpa chemnitzia (Esper) J.V. Lamouroux is a species with great phenotypic plasticity, and its characters may overlap with those of Caulerpa racemosa (Forsskål) J. Agardh (Belton et al., 2014), which is a very common and widespread species on the coast of Colombia. Patiño and Flórez (1993) reported C. chemnitzia for the Gulf of Morrosquillo; however, there is no herbarium material to corroborate the presence of the species, and it is considered a doubtful record. Future sampling campaigns could resume the search for this species in the localities previously reported by Patiño and Flórez (1993), such as the beaches of San Antero (Córdoba) and the mangrove ecosystems of Coveñas, Santiago de Tolú and San Onofre, in the department of Sucre.
The species Polysiphonia subtilissima Montagne has been reported on the basis of field photographs. Species in the genus Polysiphonia Greville are separated on the basis of the number of pericentral cells, type of connection between rhizoids and pericentral cells, and origin of branches, characters which are not distinguishable in the field and mostly require mounting and observation with an optical microscope (Stuercke and Freshwater, 2008). Therefore, in the present work the taxon is considered a doubtful record. Nevertheless, the complexity of the taxonomy of the genus Polysiphonia denotes the need to implement molecular tools to detect more species of the genus that are potentially inhabiting the Gulf of Morrosquillo and other regions of the Colombian Caribbean, as has already been done with other species of Polysiphonia in the Panamanian Caribbean (Mamoozadeh and Freshwater, 2012) and other regions of the world (Kim and Kim, 2014; Díaz-Tapia et al., 2020). In the Colombian context, it is essential to address the mentioned problem, not only to broaden the knowledge of the diversity of macroalgae in the Colombian Caribbean, but also because of the cryptic nature of Polysiphonia (Geoffroy et al., 2012; Hu and Fraser, 2016; Díaz-Tapia et al., 2020) and its classification as the world's most invasive non-native algae genus (Thomsen et al., 2016). The latest report by Wynne (2022), which indicates the presence of 27 species of the genus (including native and foreign species) in the tropical and subtropical Western Atlantic Ocean, is a wake-up call for the environmental authorities of the country to generate local policies on the knowledge, prevention and control of invasive algae, which eventually, as indicated by Thomsen et al. (2016), can drive changes in the structure of marine biological communities. Based on current knowledge, it is not possible to affirm that Polysiphonia subtilissima or other Polysiphonia species represent a management problem (invasion) for the coasts of the Gulf of Morrosquillo, but it does show the need to increase the sampling effort and the investment of public and private funds that promote more specific research that helps to measure the impact that some of these species (and others) may have on the communities of the estuary and coral ecosystems of the region.
Considering that at the moment the macroalgae reported for the Caribbean coast of Colombia reach a number of 619 taxa (Rincón-Díaz and Gavio, 2020), and that the list of species of the tropical and subtropical Western Atlantic Ocean has recently been updated, with 1861 taxa, of which four are new names (Wynne, 2022), it is evident that the integrated report of 97 taxa in the Gulf of Morrosquillo is an indicator that shows a low knowledge of the algal diversity of this region, and we expect a large number of species yet to be recorded. However, of the total species registered for the Colombian Caribbean (619), 14% are represented in the Gulf of Morrosquillo, projecting it as a relevant ecosystem for the survival of Colombia's algal biodiversity. This potential diversity can be used by the new generations of researchers interested in Phycology, so that, together with the environmental authorities, valuable and quantitative information can be provided allowing more precise decision-making in the management, sustainable use, and marine-coastal conservation of the Colombian Caribbean.
Acknowledgements
The authors wish to thank all the people working at the libraries of the Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis (INVEMAR), Universidad de Córdoba, Universidad Jorge Tadeo Lozano, Universidad del Magdalena and Universidad de Sucre. Gloria Garduño Solórzano (Universidad Nacional Autónoma de México) and Citlali Galicia García (CVU TecNM, Mexico) are acknowledged for providing critical references. Thanks to the Centro de Investigaciones Oceanográficas e Hidrográficas del Pacífico (CIOH, Colombia), Anny Marmolejo and Christian Bermudez-Rivas for logistical support. We appreciate the support of Michael J. Wynne (University of Michigan, USA) to share information on the updated checklist of western Caribbean algae and critical review of the manuscript, and Juan Felipe Ortega Giraldo (Imaging laboratory of the postgraduate degree in Sciences-Biology of the Universidad del Valle) for taking photographs of Bostrychia tenella. We greatly appreciate the contribution of Diana N. Duque-Gamboa and Rubén D. González-Román (both at Universidad de Cundinamarca), for their valuable suggestions regarding the taxonomic comments of the manuscript. To Christian Bermudez Rivas (Pacific Oceanographic and Hydrographic Research Center) for the support in the final edition of the map.
Literature cited
Aguilera-Díaz, M. 2005. La economía del Departamento de Sucre: ganadería y sector público. Documentos de trabajo sobre economía regional y urbana. No. 63. Banco de la República. Cartagena de Indias, Colombia. 129 pp.
Albis-Salas, M. and B. Gavio. 2011. Notes on marine algae in the International Biosphere Reserve Seaflower, Caribbean Colombian I: New records of macroalgal epiphytes on the seagrass Thalassia testudinum. Botanica Marina 54(6): 537-543. DOI: https://doi.org/10.1515/BOT.2011.069
Albis-Salas, M. and B. Gavio. 2015. Notes on the marine algae of the International Biosphere Reserve Seaflower, Caribbean Colombia IV: New records of macroalgal epiphytes on the seagrass Thalassia testudinum. Boletin de Investigaciones Marinas y Costeras 44(1): 55-70. DOI: https://doi.org/10.25268/bimc.invemar.2015.44.1.20
Bastidas-Salamanca, M. and A. Ordóñez-Zúñiga. 2017. Capítulo 8. Región 7: Golfo de Morrosquillo. In: Ricaurte-Villota, C. and M. Bastidas-Salamanca (eds.). Regionalización Oceanográfica: Una visión dinámica del Caribe. Instituto de Investigaciones Marinas y Costeras José Benito Vives De Andréis (INVEMAR). Serie de Publicaciones Especiales de INVEMAR #14. Santa Marta, Colombia. Pp. 124-137.
Belton, G., W. van Reine, J. Huisman, S. Draisma and C. Gurgel. 2014. Resolving phenotypic plasticity and species designation in the morphologically challenging Caulerpa racemosa-peltata complex (Chlorophyta, Caulerpaceae). Journal of Phycology 50(1): 32-54. DOI: https://doi.org/10.1111/jpy.12132
Bula-Meyer, G. 1998. Estado actual de la taxonomía de las macroalgas marinas de Colombia. Boletín Ecotrópica, Ecosistemas Tropicales 33: 1-14.
Bula-Meyer, G. and G. Díaz-Pulido. 1995. Macroalgas del banco de las animas y nuevos registros para el Caribe colombiano. Anales del Instituto de Investigaciones Marinas de Punta de Betín 24: 173-183.
Bulan, J., S. Maneekat, G. C. Zuccarello and N. Muangmai. 2022. Phylogeographic patterns in cryptic Bostrychia tenella species (Rhodomelaceae, Rhodophyta) across the Thai-Malay Peninsula. Algae 37(2): 123-133. DOI: https://doi.org/10.4490/algae.2022.37.6.4
Bykova, N., S. T. LoDuca, Q. Ye, V. Marusin, D. Grazhdankin and S. Xiao. 2020. Seaweeds through time: Morphological and ecological analysis of Proterozoic and early Paleozoic benthic macroalgae. Precambrian Research 350: 105875. DOI: https://doi.org/10.1016/j.precamres.2020.105875
Cabarcas, M. and R. García-Ureña. 2020. Catálogo de macroalgas del banco de las ánimas. In: García-Ureña, R., A. Acero, S. Zea, N. Ardila, H. Hernández-Hamón, M. Cabarcas and E. Pineda-Munive (eds.). El banco de las ánimas: valoración de su biodiversidad. Universidad del Magdalena. Editorial Unimagdalena. Santa Marta, D.T.C.H., Colombia. Pp. 77-122.
Camacho, O. and J. Montaña-Fernández, J. 2012. Cultivo experimental en el mar del alga roja Hypnea musciformis en el área de Santa Marta, Caribe Colombiano. Boletin de Investigaciones Marinas y Costeras 41(1): 29-46. DOI: https://doi.org/10.25268/bimc.invemar.2012.41.1.71
Castaño, C. 2002. Capítulo 3: Golfos y Bahías del Caribe colombiano. In: Castaño, C. (ed.). Golfo y Bahías de Colombia. Banco de Occidente. Cali, Colombia. Pp. 1-30.
Cheney, D. 1977. R&C/P A new and and improved ratio for comparing seaweed floras. Journal of Phycology 13(S): 1-13.
Dawes, C. and A. Mathieson. 2008. The seaweeds of Florida. University Press of Florida. Gainesville, USA. 656 pp.
Daza, C. 2023. Estudio de la calidad ambiental marina a partir de un índice con el uso de algas y cianobacterias en las islas de Providencia y Santa Catalina, Reserva de la Biosfera Seaflower, Colombia. Trabajo de grado. Facultad de Ciencias Agropecuarias, Programa de Ingeniería Ambiental. Universidad de Cundinamarca, Facatativá. Cundinamarca, Colombia. 70 pp.
Díaz-Merlano, J. and M. Puyana. 1994. Moluscos del Caribe colombiano, un catálogo ilustrado. Departamento Administrativo de Ciencia, Tecnología e Innovación (Colciencias), Fundación Natura e Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis (INVEMAR). Bogotá, D.C., Colombia. 377 pp.
Díaz-Pulido, G. 2000. Vegetación marina de un sector de la plataforma continental de la Guajira (Caribe colombiano). Boletín de Investigaciones Marinas y Costeras 29: 27-34.
Díaz-Pulido, G. and G. Bula-Meyer. 1997. Marine algae from oceanic atolls in the Southwestern Caribbean (Albuquerque Cays, Courtown Cays, Serrana Bank, and Roncador Bank). Atoll Research Bulletin 448: 1-18. DOI: https://doi.org/10.5479/si.00775630.448.1
Díaz-Pulido, G. and M. Díaz-Ruíz. 2003. Diversity of benthic marine algae of the Colombian Atlantic. Biota Colombiana 4: 203-246.
Díaz-Tapia, P., M., M. Ly and H. Verbruggen. 2020. Extensive cryptic diversity in the widely distributed Polysiphonia scopulorum (Rhodomelaceae, Rhodophyta): Molecular species delimitation and morphometric analyses. Molecular Phylogenetics and Evolution 152: 106909. DOI: https://doi.org/10.1016/j.ympev.2020.106909
Florez-Leiva, L., B. Gavio, M. Díaz-Ruiz, O. Camacho and G. Díaz-Pulido. 2010. Recolección y preservación de macroalgas marinas: una guía para estudios ficológicos. Intropica: Revista del Instituto de Investigaciones Tropicales 5: 97-103.
Garnica, L. 2021. Perspectivas del ordenamiento territorial marino costero a partir de los ecosistemas estratégicos en Colombia. Trabajo de grado. Universidad de Ciencias Aplicadas y Ambientales (U.D.C.A), Facultad de Ciencias Ambientales y de la Sostenibilidad. Ingeniería Geográfica y Ambiental. Bogotá, D.C., Colombia. 90 pp.
Gavio, B. 2023. Capítulo 15: Historia de la Ficología en Colombia. In: Campos, N. (ed.). Colombia Bioazul: Dos mares un país; Territorios por explorar. Universidad Nacional de Colombia. Instituto de Estudios en Ciencias del Mar-CECIMAR. Santa Marta, Colombia. Pp. 239-253.
Gavio, B. and C. Daza. 2022. Macroalgas de las islas de Providencia y Santa Catalina. Universidad de Cundinamarca, Universidad Nacional de Colombia. Bogotá, D.C., Colombia. 12 pp.
Gavio, B., M. Cifuentes-Ossa and M. Wynne. 2015. Notes on the marine algae of the International Biosphere Reserve Seaflower, Caribbean Colombia V: First study of the algal flora of Quitasueño Bank. Boletin de Investigaciones Marinas y Costeras 44(1): 117-126. DOI: https://doi.org/10.25268/bimc.invemar.2015.44.1.23
Gavio, B., J. Prato, M. Gnecco, M. F. Maya and J. E. Mancera-Pineda. 2022a. Bloom of Trichogloeopsis pedicellata (Rhodophyta, Nemaliales) following hurricane Iota in San Andrés, Southwestern Caribbean Sea. Frontiers in Marine Science 9: 1-7. DOI: https://doi.org/10.3389/fmars.2022.978869
Gavio, B., N. Rincón-Díaz, L. Chasqui, M. Luna and M. Puyana. 2022b. Spatial variation of vegetative growth of the introduced species Caulerpa scalpelliformis along La Guajira Peninsula, Colombia. Nova Hedwigia 115(3-4): 307-316. DOI: https://doi.org/10.1127/nova_hedwigia/2022/0717
Geoffroy, A., L. Le Gall and C. Destombe. 2012. Cryptic introduction of the red alga Polysiphonia morrowii Harvey (Rhodomelaceae, Rhodophyta) in the North Atlantic Ocean highlighted by a DNA barcoding approach. Aquatic Botany 100: 67-71. DOI: https://doi.org/10.1016/j.aquabot.2012.03.002
Gómez-Cubillos, M. 2018. Interacciones coral-césped de algas y sus efectos en la comunidad coralina en el área de Santa Marta, Caribe colombiano. Trabajo de investigación maestría. Universidad Nacional de Colombia, Instituto de Estudios en Ciencias del Mar (CECIMAR), Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis (INVEMAR). Santa Martha, D.T.C.H, Colombia.
Goreau, T. 1963. Calcium carbonate deposition by coralline algae and corals in relation to their roles as reef-builders. Annals of the New York Academy of Sciences 109(1): 127-167. DOI: https://doi.org/10.1111/j.1749-6632.1963.tb13465.x
Guiry, M. and G. Guiry. 2024. Algae base: World-wide electronic publication. National University of Ireland. Galway, Ireland. https://www.algaebase.org (consulted February, 2024).
Guiry, M., Ø. Moestrup and R. Andersen. 2023. Validation of the phylum name Heterokontophyta. Notulae Algarum 297: 1-5.
Hernández-Contreras, D. 2017. Taxonomía y sistemática molecular del género Bostrychia (Rhodophyta: Ceramiales) en el Pacífico colombiano. Trabajo de investigación Maestría en Ciencias-Biología. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Cali, Valle del Cauca, Colombia. 93 pp.
Hu, Z. and C. Fraser. 2016. Seaweed phylogeography: Adaptation and evolution of seaweeds under environmental change. Springer. Dordrecht, Netherlands. DOI: https://doi.org/10.1007/978-94-017-7534-2
Hurtado, A., I. Neish and A. Critchley. 2019. Phyconomy: the extensive cultivation of seaweeds, their sustainability and economic value, with particular reference to important lessons to be learned and transferred from the practice of eucheumatoid farming. Phycologia 58(5): 472-483. DOI: https://doi.org/10.1080/00318884.2019.1625632
ITIS. 2024. Integrated Taxonomic Information System (ITIS) on-line database. https://www.itis.gov/ (consulted february, 2024).
Kim, B. and M. Kim. 2014. Three new species of Polysiphonia sensu lato (Rhodophyta) based on the morphology and molecular evidence. Algae 29(3): 183-195. DOI: https://doi.org/10.4490/algae.2014.29.3.183
Kim, G. and I. Lee. 1990. A taxonomic reappraisal of Antithamnionella breviramosa (Dawson) Wollaston (Rhodophyta, Ceramiaceae). The Korean Journal of Phycology 5(2): 117-122.
Littler, D. and M. Littler. 2000. Caribbean reef plants. An identification guide to the reef plants of the Caribbean, Bahamas, Florida and Gulf of Mexico. OffShore Graphics, Inc. Washington, D.C., USA. 542 pp.
Littler, D., M. Littler and D. Hanisak. 2008. Submersed plants of the Indian River Lagoon. A floristic inventory and field guide. OffShore Graphics, Inc. Washington, D.C., USA. 286 pp.
López-Jiménez, I., L. Quan-Young and L. Florez-Leiva. 2021. Effect of terrigenous sediments on macroalgae functional-form groups of coral reefs in Capurganá, Colombian Caribbean. Scientia Marina 85(2): 125-135. DOI: https://doi.org/10.3989/scimar.05146.013
Macroalgal Herbarium Portal. 2024. Macroalgal Herbarium Portal. https://macroalgae.org/portal/index.php (consulted february, 2024).
Mamoozadeh, N. and D. Freshwater. 2012. Polysiphonia sensu lato (Ceramiales, Florideophyceae) species of Caribbean Panama including Polysiphonia lobophoralis sp. nov. and Polysiphonia nuda sp. nov. Botanica Marina 55(4): 317-347. DOI: https://doi.org/10.1515/bot-2012-0147
Mateo-Cid, L., A. Mendoza-González, R. Alvarado-Villanueva, M. Ortega-Murillo, J. Ceballos-Corona, J. Sánchez-Heredia and I. Méndez-Guzmán. 2020. Lista actualizada y nuevos registros de las algas rojas (Rhodophyta) del litoral de Michoacán, México. Revista Mexicana de Biodiversidad 91: 1-22. DOI: https://doi.org/10.22201/IB.20078706E.2020.91.3353
Mathieson, A., C. Dawes, E. Hehre and L. Harris. 2009. Floristic studies of seaweeds from Cobscook Bay, Maine. Northeastern Naturalist 16(5): 1-48. DOI: https://doi.org/10.1656/045.016.m501
Mendoza-Castro, Y. and A. Suarez-Acuña. 2018. Macroalgas Chlorophyceae asociadas al litoral rocoso del Cabo de la Vela, Caribe colombiano. Ciencia e Ingeniería 5(1): 12-21.
Meneses, D. and J. Rivera. 2014. Evaluación química del extracto total de Hypnea valentiae y potencial actividad larvicida en Aedes aegypti. Trabajo de grado. Facultad de Educación y Ciencias, Universidad de Sucre. Sincelejo, Sucre, Colombia. 79 pp.
Neiva, J., E. Serrão, J. Assis, G. A. Pearson, J. A. Coyer, J. L. Olsen, G. Hoarau and M. Valero. 2016. Part IV. Chapter 11: Climate oscillations, range shifts and phylogeographic patterns of North Atlantic Fucaceae. In: Hu, Z. and C. Fraser (eds.). Seaweed Phylogeography: Adaptation and Evolution of seaweeds under environmental change. Dordrecht, Netherlands. Pp. 279-308. DOI: https://doi.org/10.1007/978-94-017-7534-2_11
Ortega, A., N. Geraldi, I. Alam, A. Kamau, S. Acinas, R. Logares, J. Gasol, R. Massana, D. Krause-Jensen and C. Duarte. 2019. Important contribution of macroalgae to oceanic carbon sequestration. Nature Geoscience 12: 748-754. DOI: https://doi.org/10.1038/s41561-019-0421-8
Ortiz, J. and B. Gavio. 2010. Notes on the marine algae of the international biosphere reserve seaflower, Caribbean Colombia II: Diversity of drift algae in San Andres island, Caribbean Colombia. Caribbean Journal of Science 46(2-3): 313-321. DOI: https://doi.org/10.18475/cjos.v46i2.a19
Palencia, R., Y. Díaz, I. Muñoz, W. Pérez and J. Vargas. 2023. Riesgo hacia la fauna y la flora por grupos armados en Colombia. Remulci 1: 41-56.
Patiño, F. and F. Flórez. 1993. Ecología marina del golfo de Morrosquillo. Universidad Nacional de Colombia, Fondo FEN. Bogotá, D.C., Colombia. 109 pp.
Peña-Salamanca, E. 2008. Dinámica espacial y temporal de la biomasa algal asociada a las raíces de mangle en la bahía de Buenaventura, costa Pacífica de Colombia. Boletín de Investigaciones Marinas y Costeras 37(2): 55-70. DOI: https://doi.org/10.25268/bimc.invemar.2008.37.2.191
Puccini, R. and G. Molina. 2022. Aportes para monitoreo y conservación de especies coralinas en el ecosistema costero del Cabo de la Vela en la Guajira, colombiana. Ciencia e Ingeniería 9(2): 1-33.
QGIS, 2023. Geographic Information System (3.34 version). QGIS Desktop 3.34 User Guide. QGIS Project. https://www.qgis.org/es/site/
Queirós, A., N. Stephens, S. Widdicombe, K. Tait, S. McCoy, J. Ingels, S. Rühl, R. Airs, A. Beesley, G. Carnovale, P. Cazenave, S. Dashfield, E. Hua, M. Jones, P. Lindeque, C. McNeill, J. Nunes, H. Parry, C. Pascoe, C. Widdicombe, T. Smyth, A. Atkinson, D. Krause-Jensen and P. Somerfield. 2019. Connected macroalgal-sediment systems: blue carbon and food webs in the deep coastal ocean. Ecological Monographs 89(3): 1-21. DOI: https://doi.org/10.1002/ecm.1366
Quintana-M., H. and J. Mercado-Gómez. 2017. Composición de dinoflagelados epífitos y forófitos en la Costa norte del golfo de Morrosquillo, Sucre, Colombia. Revista Colombiana de Ciencia Animal 9(2): 129-140. DOI: https://doi.org/10.24188/recia.v9.n2.2017.550
Quirós-Rodríguez, J., J. Arias-Ríos and R. Ruiz. 2010. Diversity of benthic marine algae of the Colombian Atlantic. Caldasia 32: 339-354.
Quirós-Rodríguez, J., P. Ramírez and N. Campos. 2013. Poliquetos (Annelida: Polychaeta) asociados a algas rojas intermareales de Córdoba, Caribe colombiano. Revista de Biología Marina y Oceanografía 48(1): 87-98. DOI: https://doi.org/10.4067/S0718-19572013000100008
Raz, L. and H. Agudelo-Zamora. 2023. Herbario Nacional Colombiano (COL), Version 13.16. Universidad Nacional de Colombia. DOI: https://doi.org/10.15472/ea8sek
Reyes-Gómez, V., H. Velasquez and B. Gavio. 2021. Notes on the marine algae of the International Biosphere Reserve Seaflower, Caribbean Colombia VIII: new records of Red Algae (Rhodophyta) from San Andres, Old Providence, and Saint Cataline, Colombia. Acta Botanica Mexicana 128: e1848: 1-23. DOI: https://doi.org/10.21829/ABM128.2021.1848
Rincón-Díaz, M. 2014. Relación espaciotemporal entre macroalgas y peces herbívoros en un arrecife insular. Tesis de Doctorado en Ciencias, Universidad Nacional de Colombia-Sede Caribe. San Andrés Islas, Colombia. 115 pp.
Rincón-Díaz, M. and B. Gavio. 2020. Diversidad de macroalgas marinas del Caribe colombiano. Version 2.8. Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis - INVEMAR. Dataset/Checklist. DOI: https://doi.org/10.15472/alecqe
Rincón-Díaz, M. and F. Ramos. 2016. Macroalgas marinas: El universo productivo de la Reserva de la Biosfera Seaflower. In: Vides, M., D. Alonso, E. Castro and N. Bolaños (eds.). Biodiversidad del mar de los siete colores. Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis - INVEMAR y Corporación para el Desarrollo Sostenible del Archipiélago de San Andrés, Providencia y Santa Catalina - CORALINA. Serie de Publicaciones Generales del INVEMAR No. 84. Santa Marta, Colombia. Pp. 40-55.
Rincón-Díaz, M., B. Gavio, M. Wynne and A. Santos-Martínez. 2018. Notes on marine algae in the international biosphere reserve seaflower, Caribbean Colombia, VII: Additions to the benthic flora of San Andrés Island. Caldasia 40(1): 97-111. DOI: https://doi.org/10.15446/caldasia.v40n1.64597
Saengkaew, J. S. Bovonsombut and Y. Peerapornpisal. 2016. Species diversity and distribution of mangrove-associated red alga Bostrychia (Rhodomelaceae, Rhodophyta) from southern Thailand. International Journal of Applied Environmental Sciences 11(1): 55-71.
Salazar-Forero, C., B. Gavio and M. Wynne. 2021. Macroalgae associated with aerial roots of Rhizophora mangle in Islas del Rosario, Colombia, Southwestern Caribbean. Caldasia 43(1): 94-104. DOI: https://doi.org/10.15446/caldasia.v43n1.85228
Sampedro-M., A., S. Prasca-S, D. Suárez-V. and L. Escobar-S. 2012. Estado de las poblaciones de quitones (Mollusca: Polyplacophora) en rompeolas artificiales de Coveñas, Sucre, Colombia. Caldasia 34: 397-407.
Schneider, C., E. Cianciola, T. Popolizio, D. Spagnuolo and C. Lane. 2015. A molecular-assisted alpha taxonomic study of the genus Centroceras (Ceramiaceae, Rhodophyta) in Bermuda reveals two novel species. Algae 30(1): 15-33. DOI: https://doi.org/10.4490/algae.2015.30.1.015
Schnetter, R. 1969. Beitrag zur Kenntnis der Algenflora an der kolumbianischen Küste der Karibischen See. Mitteilungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas “Punta de Betín” 3: 49-57.
Schnetter, R. 1976. Marine Algen der karibischen Küsten von Kolumbien I: Phaeophyceae. Bibliotheca Phycologica 24: 1-125.
Schnetter, R. 1978. Marine Algen der karibischen Küsten von Kolumbien II: Chlorophyceae. Bibliotheca Phycologica 42: 1-199.
Stuercke, B. and D. Freshwate. 2008. Consistency of morphological characters used to delimit Polysiphonia sensu lato species (Ceramiales, Florideophyceae): Analyses of North Carolina, USA specimens. Phycologia 47(6): 541-559. DOI: https://doi.org/10.2216/08-09.1
Tano, S., M. Eggertsen, S. Wikström, C. Berkström, A. Buriyo and C. Halling. 2016. Tropical seaweed beds are important habitats for mobile invertebrate epifauna. Estuarine, Coastal and Shelf Science 183(Part A): 1-12. DOI: https://doi.org/10.1016/j.ecss.2016.10.010
Taylor, W. 1960. Marine algae of the eastern tropical and subtropical coasts of the Americas. University of Michigan Press, Ann Arbor, USA , and Ambassador Books Ltd. Toronto, Canada. 870 pp.
Thomsen, M., T. Wernberg, P. South and D. Schiel. 2016. Part III. Chapter 6: Invasive seaweeds. Non-native seaweeds drive changes in marine coastal communities around the world. In: Hu, Z. and C. Fraser (eds.). Seaweed Phylogeography: Adaptation and Evolution of seaweeds under environmental change. Dordrecht, Netherlands. Pp. 147-185.
Tuya, F., L. Png-Gonzalez, R. Riera, R. Haroun and F. Espino. 2014. Ecological structure and function differs between habitats dominated by seagrasses and green seaweeds. Marine Environmental Research 98: 1-13. DOI: https://doi.org/10.1016/j.marenvres.2014.03.015
Umanzor, S., L. Ladah, L. E. Calderon-Aguilera and J. A. Zertuche-González. 2019. Testing the relative importance of intertidal seaweeds as ecosystem engineers across tidal heights. Journal of Experimental Marine Biology and Ecology 511: 100-107. DOI: https://doi.org/10.1016/j.jembe.2018.11.008
Vandendriessche, S., M. Messiaen, S. O’Flynn, M. Vincx and S. Degraer. 2007. Hiding and feeding in floating seaweed: Floating seaweed clumps as possible refuges or feeding grounds for fishes. Estuarine, Coastal and Shelf Science 71(3-4): 691-703. DOI: https://doi.org/10.1016/j.ecss.2006.09.017
Vasquez-Carrillo, C. and K. Sullivan. 2018. Diversity and extent of coastal submerged aquatic vegetation in an unexplored coastal upwelling region of the Caribbean Sea. International Journal Biodiversity Endangered Species 2018(01): 1-13.
Vega-Sequeda, J., A. Rodríguez-Ramírez, M. Reyes-Nivia, M. and R. Navas-Camacho. 2008. Formaciones coralinas del área de Santa Marta: Estado y patrones de distribución espacial de la comunidad bentónica. Boletin de Investigaciones Marinas y Costeras 37(2): 87-105. DOI: https://doi.org/10.25268/bimc.invemar.2008.37.2.193
Verborgh, R. and M. De Wilde. 2013. Using OpenRefine. Packt Publishing Ltd. Birmingham, UK.
Won, B., T. Cho and S. Fredericq. 2009. Morphological and molecular characterization of species of the genus Centroceras (Ceramiaceae, Ceramiales), including two new species. Journal of Phycology 45(1): 227-250. DOI: https://doi.org/10.1111/j.1529-8817.2008.00620.x
WoRMS. 2023. World Register of Marine Species. https://www.marinespecies.org/ (consulted November, 2023).
Wynne, M. 2022. Checklist of benthic marine algae of the tropical and subtropical Western Atlantic: fifth revision. Nova Hedwigia Beiheft 153: 1-180.
Zerrifi, S., F. Khalloufi, B. Oudra and V. Vasconcelos. 2018. Seaweed bioactive compounds against pathogens and microalgae: Potential uses on pharmacology and harmful algae bloom control. Marine Drugs 16(2): 55. DOI: https://doi.org/10.3390/md16020055
Zuccarello, G. and J. West. 2011. Insights into evolution and speciation in the red alga Bostrychia: 15 years of research. Algae 26(1): 3-20. DOI: https://doi.org/10.4490/algae.2011.26.1.003
Zuccarello, G., N. Muangmai, M. Preuss, L. Sanchez, S. L. de Goër and J. West. 2015. The Bostrychia tenella species complex: morphospecies and genetic cryptic species with resurrection of B. binderi. Phycologia 54(3): 261-270. DOI: https://doi.org/10.2216/15-005.1
Appendix

Author contributions
Funding
Author notes
4Author for correspondence: humberto19quintana@gmail.com