Articles
A positively invariant attracting set for a predator-prey model with a non-differentiable functional response
Un conjunto atractor positivamente invariante para un modelo depredador-presa con respuesta funcional no-diferenciable
A positively invariant attracting set for a predator-prey model with a non-differentiable functional response
Selecciones Matemáticas, vol. 9, núm. 02, pp. 234-242, 2022
Universidad Nacional de Trujillo

Esta obra está bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Recepción: Julio , 20, 2022
Aprobación: Noviembre , 13, 2022
Abstract: In this work, the ecologically well-posedness of a predator-prey model is demonstrated by showing that a region of the first quadrant is a positively invariant attracting set for the solutions of that model. The predator-prey model considers logistic-type growth in both populations and a non-differentiable functional response that generalizes previous ones. Due to non-differentiability, there is no uniqueness of solutions, and the standard methodology cannot be applied directly. Topological equivalences, geometrical arguments, and the Poincare-Bendixson theorem are used to obtain our result.
Keywords: Predator-prey model, invariant set, functional response.
Resumen: En el presente trabajo demostramos que un modelo depredador-presa está bien puesto en el sentido ecológico a través de evidenciar que existe un conjunto atractor positivamente invariante para las soluciones del modelo. El modelo depredador-presa en estudio considera crecimiento logístico en ambas poblaciones y una respuesta funcional no-diferenciable. Esta respuesta funcional es una generalización de otras consideradas en anteriores trabajos. Por causa de la no-diferenciabilidad, no podemos garantizar unicidad de soluciones y los métodos clásicos no pueden ser aplicados directamente. Usando equivalencias topológicas, argumentos geométricos y el teorema de Poincaré-Bendixon obtenemos nuestro resultado.
Palabras clave: Modelo depredador-presa, conjunto invariante, respuesta funcional.
1. Introduction.
For the well-posedness of a mathematical model in ecology, we must ensure that the size of each population involved in it is always a non-negative number that can be increased to a certain limit. Usually, the non-negativity and boundedness of the functions that represent the population sizes are proved. This work aims to prove that there exists a positively invariant attracting set for the predator-prey model described by the following system of ordinary differential equations.
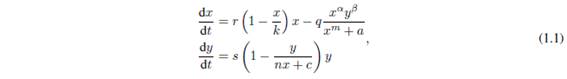
where x (t) and y (t) represent respectively the prey and predator population size at the positive time t, and with ecological parameters (see Table 1.1) satisfying
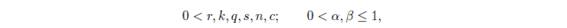
and
m=a=0 or 0 < m,a.
More precisely, we prove that any solution of system (1.1) with initial condition in the interior of the first quadrant enters in the region
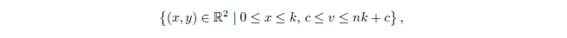
and remains in it.
System (1.1) considers the following: the predator is generalist with logistic-type growth equation in which the environmental carrying capacity is proportional to the prey population size plus a positive constant that represents an alternative food; the prey natural growth is given by a logistic equation; the action of the predator consuming the prey, called functional response, is represented by the function 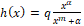 , which is a generalization of the Rosenzweig functional response [1] and describes anti-predator behaviors (prey herd behavior, group defense formation); the depredation rate is given by
, which is a generalization of the Rosenzweig functional response [1] and describes anti-predator behaviors (prey herd behavior, group defense formation); the depredation rate is given by 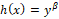 which is a modification of the Cobb-Douglas type function [2, 3] and describes predator mutual interference [4, 5].
which is a modification of the Cobb-Douglas type function [2, 3] and describes predator mutual interference [4, 5].
We point out that the logistic-type growth of predators is in fact a modified Leslie-Gower scheme [6, 7].
Note that for 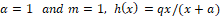 is the Holling type II functional response.
is the Holling type II functional response.
System (1.1) with m = . = 0 was studied in the past [8, 9]. The dynamics of the general case 0 < m; was not analyzed. Therefore, our result is helpful to continue with the research on predator-prey dynamics. .
System (1.1) is defined on . But, in general, (1.1) is not differentiable on the coordinate axes. For instance, when we consider 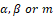 less than 1. Therefore the analysis must be careful. We can expect uniqueness of solutions only in the interior of the first quadrant. The equilibrium points of (1.1) are: (0; 0), (k; 0), and (0; c) over the coordinate axes, and (xe; ye) satisfaying the isocline equations
less than 1. Therefore the analysis must be careful. We can expect uniqueness of solutions only in the interior of the first quadrant. The equilibrium points of (1.1) are: (0; 0), (k; 0), and (0; c) over the coordinate axes, and (xe; ye) satisfaying the isocline equations
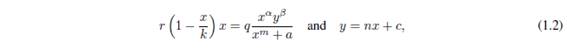
in the interior of the first quadrant. In fact, the above equations can have several solutions. Moreover, it is clear that
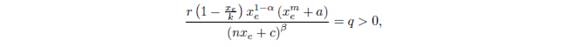
and therefore xe must belong to ]0; k[. Consequently, ye must belong to ]c; nk + c[.
The paper is organized as follows. In Section 2, we begin by proving the non-negativity and boundedness of solutions. Next, a differentiable system, topologically equivalent to (1.1), is obtained. The nature of the boundary equilibrium points is determined in section 3. In Section 4, our result follows from the application of the Poincare-Bendixson theorem and some geometrical arguments over the topologically equivalent system.
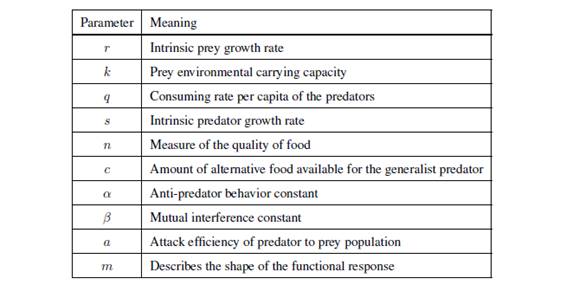
Table 1.1: The parameters of the system (1.1) and their ecological meanings.
2. Boundedness and topological equivalences.
System (1.1) describes the interaction between prey and predators. hence, the solutions of this system must be bounded and non-negative. These properties are guaranteed by the following proposition.


In order to simplify the calculations, we reduce system (1.1) to a polynomial form making a change of variable and a time rescaling. We show this in our next lemma.


Although system (1.1) is not differentiable, it is possible to obtain an equivalent differentiable system.
The equivalence is established inside the first quadrant.

Proof: System (2.2) and (2.3) are obtained by applying to system (2.1) (which is topologically equivalent on  to (1.1) by Lemma 2.1) the change of coordinates and the time rescaling given by the diffeomorphism
to (1.1) by Lemma 2.1) the change of coordinates and the time rescaling given by the diffeomorphism

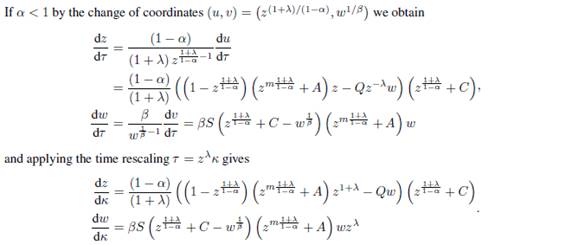
The Jacobian matrix of the diffeomorphism 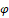 is
is
3. Equlibrium of boundary points.
Here we analyze the nature of equilibrium points over the coordinate axes.

and therefore the equilibrium point (0, 0) is an unstable node. On the other hand, note that if 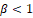 then the partial derivative of F at the equilibrium point (k, 0)
then the partial derivative of F at the equilibrium point (k, 0)
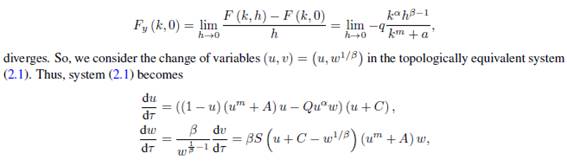
and the equilibrium point (k, 0) in the new coordinates is (1,0). If we denote by M(u,v) and N(u,v) the functions on the right side of the above equations, that is, 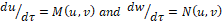 then we have
then we have
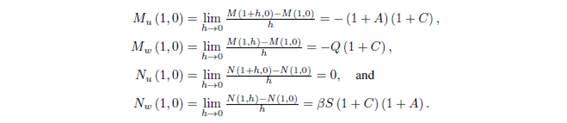
It follows that the Jacobian matrix at (1; 0) is
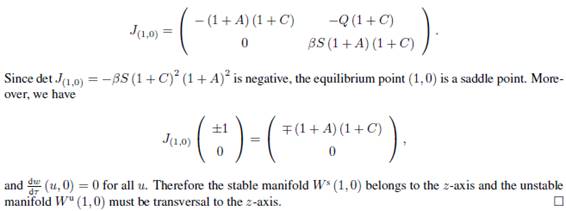
4. The positively invariant attracting set.
Our main result is the following.


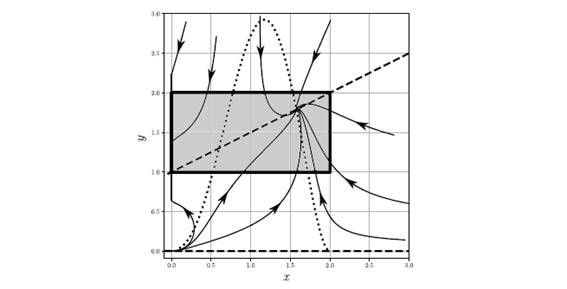
Figure 4.1: Numerical solutions of system (1.1) with parameter r=1.5, q=0.5, k=2, s=1, n=0.5, c = 1, 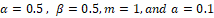 . The set A is given by the gray rectangle. The isoclines are in dotted and dashed lines.
. The set A is given by the gray rectangle. The isoclines are in dotted and dashed lines.



it follows that

5. Conclusions.
In this work, we have obtained a positively invariant attracting set for a predator-prey model that considers a non-differentiable functional response, which is a generalization of the Rosenzweig functional response [1].
In order to simplify the calcualations, a reparameterization and a time rescaling were made, obtaining the topologically equivalent system (2.1). Using this, we proved that our original system is topollogically equivalent (on the interior of the first quadrant) to a new differentiable system ((2.2) for and (2.3) for . Also, the nature of boundary equilibrium points was explored. Finally, the Poincaré-Bendixon theorem and geometrical arguments were applied to the differentiable system (2.2)-(2.3) to obtain our main result. Two cases were examined separately, 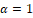 and
and 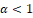 With our result, we have demonstrated the ecologically well-posedness of system (1.1).
With our result, we have demonstrated the ecologically well-posedness of system (1.1).
Although (1.1) was studied for particular parameter values [8, 9], the dynamics of the general case remains unexplored. Therefore, our result represents a first step in the analysis of predator-prey system(1.1).
6. Acknowledgements.
My deepest gratitude goes to Prof. Eduardo González-Olivares for his friendship, guidance, and scientific advice throughout the years.
References
[1] Rosenzweig ML. Paradox of enrichment: destabilization of exploitation ecosystems in ecological time. Science.
[2] Cobb CW, Douglas PH. A theory of production. Papers and Proccedings of the Fortieth Annual Meeting of the American Economic Association. 1928;18(1):139–165.
[3] Clark CW. Mathematical Bioeconomics: The Optimal Management of Renewable Resources (2nd edition). Wiley, New York; 1990.
[4] Freedman H. Stability analysis of a predator–prey system with mutual interference and density–dependent death rates. Bulletin of Mathematical Biology. 1979;41(1):67–78.
[5] González-Olivares E, Sáez E, Stange E, Szanto I. Topological Description of a Non-Differentiable Bioeconomics Model. Rocky Mountain Journal of Mathematics. 2005 08; 35:1133–1155.
[6] Leslie P, Gower J. The properties of a stochastic model for the predator-prey type of interaction between two species. Biometrika. 1960;47(3/4):219–234.
[7] Aziz-Alaoui M, Okiye MD. Boundedness and global stability for a predator-prey model with modified Leslie–Gower and Holling–type II schemes. Applied Mathematics Letters. 2003;16(7):1069–1075.
[8] Díaz-Avalos JD, González-Olivares E. A class of predator-prey models with a non-differentiable functional response. In: Vigo-Aguiar J, editor. Proceedings of the 17th International Conference on Computational and Mathematical Methods in Science and Engineering, CMMSE 2017. vol. 3; 2017. p. 765–776.
[9] Rivera-Estay V, González-Olivares E, Rojas-Palma A, Vilches-Ponce K. Dynamics of a class of Leslie-Gower predation models with a non-differentiable functional response. In: Dutta H, Peters J, editors. Applied Mathematical Analysis: Theory, Methods, and Applications, Studies in Systems, Decision and Control. vol. 117; 2020. p. 433–457.
Notes
Díaz-Avalos J. A positively invariant attracting set for a predator-prey model with a non-differentiable functional response. Selecciones Matemáticas. 2022;9(2):234–242. http://dx.doi.org/10.17268/sel.mat.2022.02.02