CARACTERIZACIÓN DE POBLACIONES DE PASTO BANDERITA (Bouteloua curtipendula) DE MÉXICO PARA SELECCIONAR GENOTIPOS CON POTENCIAL PARA PRODUCCIÓN DE SEMILLA
CHARACTERIZATION OF SIDEOATS GRAMA (Bouteloua curtipendula) POPULATIONS FROM MEXICO TO SELECT GENOTYPES WITH POTENTIAL FOR SEED PRODUCTION
CARACTERIZACIÓN DE POBLACIONES DE PASTO BANDERITA (Bouteloua curtipendula) DE MÉXICO PARA SELECCIONAR GENOTIPOS CON POTENCIAL PARA PRODUCCIÓN DE SEMILLA
Revista Fitotecnia Mexicana, vol. 40, núm. 3, pp. 309-316, 2017
Sociedad Mexicana de Fitogenética, A.C.
Resumen: La escasez de semilla de pastos forrajeros en el norte de México es una limitante para emprender programas de recuperación de pastizales. Por lo anterior, es importante considerar la diversidad y evaluar el potencial de producción de semilla que presentan algunas especies nativas apomícticas como el pasto banderita [Bouteloua curtipendula (Michx.) Torr.] en poblaciones naturales. El objetivo de este estudio fue caracterizar la diversidad morfológica de la inflorescencia de poblaciones de pasto Banderita en México, para seleccionar genotipos con potencial para producción de semilla. Se recolectaron y establecieron ex situ 168 genotipos procedentes de 10 estados de la República Mexicana. Se evaluó la morfología de la inflorescencia durante la etapa de madurez fisiológica de la semilla con los descriptores: longitud de inflorescencia (LIN), ramillas por inflorescencia (RPI), espiguillas por ramilla (EPR), longitud de ramilla (LRM), longitud de raquis (LRQ), longitud de pedicelo (LPD) y peso de semilla (PSEM). Las variables ambientales de procedencia de los genotipos se clasificaron en tres categorías (alta, media y baja) de altitud (ALT) y precipitación media anual (PPma). Los datos se analizaron mediante técnicas de análisis multivariado usando el paquete SAS. Se encontró variabilidad morfológica entre grupos de genotipos. Los tres primeros componentes principales (CP) explicaron el 72 % de la variación total observada. Los pseudo estadísticos F y T2 sugieren la formación de cuatro grupos (G) con un R2 = 0.76 y con diferenciación multivariada (P < 0.001) entre sí. La mayoría de los grupos se asociaron a las categorías media de ALT y PPma. Se encontró variabilidad y diferenciación de genotipos con alto potencial para producción de semilla, donde sobresale el G1 para esta característica. En México se dispone de riqueza genética de pasto Banderita para producción de semilla. Los genotipos que mostraron el mayor potencial para producción de semilla fueron: 1 y 7 originarios de Aguascalientes, 84 de Jalisco, y 508 y 512 de Chihuahua.
Palabras clave: Pasto nativo, inflorescencia, variabilidad morfológica.
Abstract: The shortage of forage grasses seed in Northern Mexico is a constraint to undertake grasslands restoration programs. Thus, it is important to consider the diversity and to evaluate the seed production potential of some native and apomictic species such as sideoats gramma grass [Boutelous curtipendula (Michx.) Torr.] in natural populations. The aim of this study was to characterize the inflorescence morphological diversity of the inflorescence of sideoats grama Mexican populations, to select genotypes with potential for seed production. One hundred-sixty four genotypes were collected in 10 Mexican states and then were established ex situ. The inflorescence morphology was evaluated at physiological maturity stage through the following descriptors: inflorescence length (INL), twigs per inflorescence (CPI), spikelets per twig (SPT), twigs length (TL), rachis length (RQL), pedicel length (PDL) and seed weight (SEW) were evaluated to characterize the inflorescence morphology at the reproductive stage. The prevailing environmental variables at the ecotype’s origin were classified into three categories (high, medium, and low) of altitude (ALT) and precipitation (PP). Data were analyzed through multivariate analysis techniques using the SAS software. Morphological variability among groups of genotypes was found. The first three principal components (PC) explained 72 % of the total observed variation. The statistics pseudo F and T2 suggest the formation of four groups (R2 = 0.76) with multivariate significant differences (P < 0.001) between each other. Most of the groups were associated to the values of medium category of PP and ALT. Variability and differentiation among genotypes with high potential for seed production were found, where the group GI presented the highest potential for seed production. In Mexico genetic variation of sideoats grama is available for seed production. Genotypes with the highest potential for seed production were: 1 and 7 originating from Aguascalientes, 84 from Jalisco and 508 and 512 from Chihuahua.
Keywords: Native grass, inflorescence, morphological variability.
INTRODUCCIÓN
La escasa disponibilidad de semilla de pastos forrajeros es una de las principales limitantes para la restauración de pastizales degradados en zonas áridas y semiáridas de México. En este sentido, es importante integrar la mayor diversidad posible de poblaciones naturales para evaluar el potencial de producción de semilla en especies nativas como el pasto banderita [Bouteloua curtipendula (Michx.) Torr.]. La mayoría de la semilla forrajera disponible en el mercado es importada, lo cual genera un alto costo y limita los programas de restauración (Martín et al., 2016). Otra desventaja de la semilla importada es que las variedades no siempre están adaptadas a las condiciones de México (Corrales et al., 2016). La principal limitante para atender estos ecosistemas degradados es la disponibilidad, calidad y precio de la semilla de especies forrajeras, debido a que cerca del 85 % es importada principalmente de los Estados Unidos de América a un costo elevado; por lo tanto, utilizar genotipos locales puede aumentar la probabilidad de éxito en programas de rehabilitación de pastizales (Beltrán et al., 2013; Rowe et al., 2012; Whalley et al., 2013).
No obstante, antes de utilizar estos materiales es importante realizar programas de evaluación y seleccionar los más adaptados y con mayor potencial productivo. Esto puede realizarse mediante la recolección y caracterización de genotipos de poblaciones naturales, donde se concentre la mayor diversidad de la especie de interés, distribuida en su centro de origen. La selección de genotipos se efectúa mediante la evaluación de descriptores que se relacionen con características de interés (Ferdinandez y Coulman, 2004; Morales et al., 2007); sin embargo, el sobrepastoreo ha ocasionado la pérdida de especies forrajeras nativas, problema actual en los agostaderos de México, ya que se estima que cerca del 90 % se encuentran en condición que va de regular a pobre (Jurado-Guerra et al., 2012; PACP-Ch, 2011).
El pasto banderita es una especie nativa del norte de México, que por sus características productivas y nutricionales es considera la segunda gramínea nativa de mayor importancia y más utilizada en programas de resiembra de pastizales en el norte de México, ya que se adapta a una amplia gama de eco-regiones. Al respecto, existe investigación donde se ha caracterizado la diversidad de esta especie por atributos forrajeros (Morales-Nieto et al. 2008); no obstante, se desconoce la variabilidad con respecto a componentes de la inflorescencia y el potencial de producción de semilla de poblaciones naturales de pasto banderita en México.
En este sentido, el producir semilla de especies forrajeras previamente evaluadas y seleccionadas garantiza la obtención de semilla de mayor calidad. Esto puede facilitar el establecimiento, persistencia y adaptación a las condiciones de la región (Morales y Melgoza, 2010). La presente investigación se planteó con el objetivo de caracterizar la diversidad morfológica de la inflorescencia de poblaciones de pasto banderita en México, para seleccionar genotipos con potencial para producción de semilla, bajo la hipótesis de que las poblaciones y genotipos de pasto Banderita en México presentan un comportamiento variable en su potencial para producción de semilla.
MATERIALES Y MÉTODOS
Colecta de material genético
Durante los años 2002 y 2003 se recolectaron 577 genotipos (individuo o planta perteneciente a una población) de pasto banderita en 13 estados de la República Mexicana: Aguascalientes, Chihuahua, Coahuila, Durango, Guanajuato, Jalisco, México, Oaxaca, San Luis Potosí, Sonora, Zacatecas, Querétaro y Guerrero. El criterio para realizar la recolecta y buscar diversidad dentro de la especie fue que cada población estuviera distante entre 50 y 100 km. Se recolectaron plantas de aproximadamente una pulgada de diámetro, provistos de raíz y biomasa aérea. Se obtuvieron de cuatro a cinco plantas por población y la biomasa aérea se podó entre 15 y 20 cm de altura. Para facilitar su transporte, el material se aisló con cinta adhesiva y se asignó un número de identificación. Los genotipos se colocaron en cajas de plástico, provistas con suelo y sustrato Peat Moss® y Agrolita® en proporción 3:1, humedecidos para su trasplante. Se obtuvieron los siguientes datos de origen: número de genotipo, sitio de recolecta, coordenadas geográficas, precipitación media anual (PPma), altitud (ALT), temperatura media anual (TEMP), nombre del recolector, tipo de asociación con otras especies y otras observaciones relacionadas con el fenotipo.
Trasplante y manejo del material
Los pastos fueron trasplantados entre junio y julio en los años de la colecta y conservados ex situ en el área agronómica del Colegio de Postgraduados, Campus Salinas de Hidalgo, San Luis Potosí, México. El terreno presentó una pendiente de 0.5 a 2.0 % y suelo vertisol con textura arcillosa (Quero-Carrillo et al., 2016). Este sitio presenta temperatura media anual de 17.1 ºC, PPma de 350 mm y altitud de 2070 msnm (INEGI, 2002). La preparación del área se realizó mediante barbecho, rastreo y nivelación; además, se cuadriculó el terreno en distancia de 1 x 1 m; en cada punto de intersección se prepararon cavidades de 50 cm de profundidad por 25 cm de diámetro para el trasplante. Posteriormente, los genotipos fueron trasplantados bajo un diseño completamente al azar; con una planta como caso o entidad por genotipo.
Para facilitar la supervivencia del material trasplantado, se aplicaron 2 L de agua en cada cavidad antes del trasplante; posterior a ello el manejo fue bajo condiciones de temporal. El área de establecimiento fue protegida con alambre de púas y malla gallinera. De los 577 genotipos trasplantados, al tercer año sobrevivieron 311; sin embargo, por ser algunos originarios del mismo sitio, sólo se evaluaron 168 genotipos procedentes de 10 estados de la República Mexicana.
Variables evaluadas
La caracterización morfológica se realizó en octubre de 2005, durante la etapa de madurez fisiológica de la semilla. Se calificaron los siguientes descriptores (variables) relacionados con producción de semilla: longitud de inflorescencia (LIN), ramillas por inflorescencia (RPI), número de espiguillas por ramilla (EPR), longitud de ramillas en mm (LRM), longitud del raquis de la inflorescencia (LRQ) en cm, longitud del pedicelo (LPD) en cm y peso de semilla por genotipo (PSEM) en g.
Análisis estadístico
El programa estadístico utilizado fue SAS 9.1.3 (SAS Institute, 2006) y el análisis de los datos fue soportado con diferentes técnicas de análisis multivariado. Se inició con análisis de correlación para identificar tipo y grado de la asociación lineal entre pares de variables, posteriormente se realizó análisis de componentes principales a partir de la matriz de correlación. Además se realizó un análisis de agrupamiento con el método de ligamiento de Ward. El número de grupos se determinó con base en el coeficiente de determinación de los pseudo estadísticos F y T2 y se analizó la significancia de la agrupación sugerida mediante análisis multivariante de la varianza (sintaxis MANOVA). Se utilizaron contrastes ortogonales para determinar diferencias entre los grupos generados por el análisis de conglomerados. Finalmente se llevó a cabo un análisis de correspondencias donde se fijó un nivel de significancia (α) de 0.1, debido a la variabilidad de las condiciones ambientales de los sitios de origen de los genotipos evaluados. En este análisis se evaluaron las relaciones entre niveles categóricos de altitud (ALT) y precipitación media anual (PPma) que presentó el lugar de origen de cada genotipo recolectado, con los grupos conformados en el análisis de agrupamiento. Los rangos establecidos para los grupos fueron: 800 a 1400, 1401 a 2000 y ≥ 2001 msnm para ALT y 100 a 350, 351 a 450 y ≥ 451 mm para PPma.
Para presentar la distribución geográfica de los grupos conformados se realizó un mapa de distribución mediante el programa ArcGIS v.9.3.
RESULTADOS Y DISCUSIÓN
Análisis de correlación
La mayoría de las variables morfológicas mostraron una correlación significativa (P ≤ 0.001). La variable LIN se correlacionó positivamente con todas las variables medidas (P ≤ 0.001); no obstante, la más fuerte asociación lineal se presentó con RPI (r = 0.72). Esto indica que al encontrar una inflorescencia de mayor longitud en pasto banderita se obtendrá mayor cantidad de ramillas por inflorescencia. Las variables que no presentaron asociación lineal con RPI fueron LRM y LPD (P > 0.05), lo que indica que la longitud de ramillas y pedicelo no influyeron sobre la cantidad de ramillas por inflorescencia.
Aunque el peso de semilla mostró una correlación significativa con algunas variables (como LIN; r = 0.29; P ≤ 0.0001) la asociación fue baja. Esto indica que la producción de semilla es una variable de comportamiento eminentemente multivariado en sus causas; es decir, que su expresión está en función de varios factores, algunos no necesariamente morfométricos; por ejemplo, en torno a su madurez, la semilla es muy irregular aun en la inflorescencia y depende del grado de dehiscencia de la especie. Por lo anterior, es importante evaluar características de la inflorescencia para estimar el potencial de la especie para producir semilla.
Análisis de componentes principales
En el análisis de componentes principales (ACP) el Cuadro 1 muestra los valores propios y la contribución a la varianza total de los componentes principales. Se encontró que los primeros componentes principales explicaron el 72 % de la variabilidad total (43 % para CP1, 17 % para CP2 y 12 % para CP3). En otros estudios, donde se caracterizó la diversidad morfológica de zacate punta blanca (Digitaria californica) y gigante (Leptochloa dubia) se explicó el 79 y 75 % de la variación total con tres componentes principales, respectivamente (Morales et al., 2012; Morales-Nieto et al., 2013). Esto indica que las poblaciones de especies nativas en México presentan amplia diversidad. En contraste, Schellenberg et al. (2012) al evaluar el comportamiento en la variación fenotípica de colecciones de pasto banderita recolectadas en diversas localidades de Canadá, reportaron que los dos primeros CP explicaron hasta en 91 % la variación total.
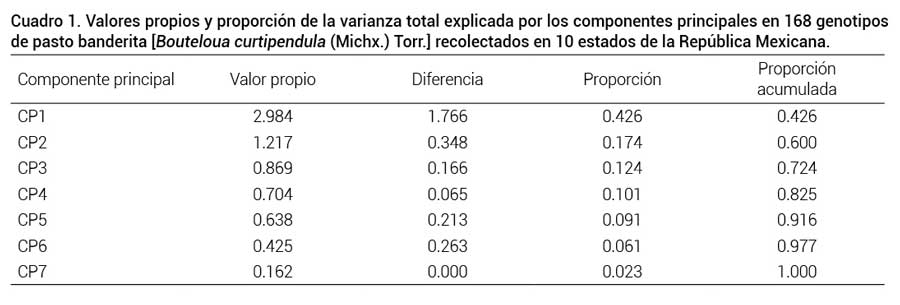
Valores propios y proporción de la varianza total explicada por los componentes principales en 168 genotipos de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recolectados en 10 estados de la República Mexicana.
En el Cuadro 2 se observan los vectores propios y la proporción con que contribuye cada variable al componente derivado. Para el CP1 las variables de mayor aporte fueron LIN, EPR y LRM, en el CP2 fueron RPI y LRM las que aportaron en mayor proporción. Para constituir el CP3, fue PSEM la variable con mayor aportación. En diversas investigaciones realizadas en especies del genero Bouteloua se concluye que la evaluación de la variabilidad morfológica constituye la base para seleccionar materiales sobresalientes para diferentes propósitos (Morales-Nieto et al., 2008; Morales et al., 2009). Una ventaja del pasto banderita es que éste se reproduce en forma apomíctica (asexual); por lo tanto, al encontrar un genotipo con características deseables, generaciones futuras tendrán consecutivamente la misma expresión fenotípica y composición genotípica (Morales-Nieto et al., 2008).
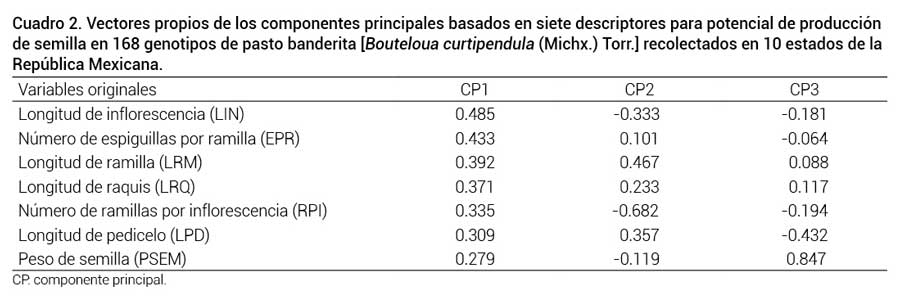
Vectores propios de los componentes principales basados en siete descriptores para potencial de producción de semilla en 168 genotipos de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recolectados en 10 estados de la República Mexicana.
CP: componente principal.Al correlacionar los primeros dos CP con las variables originales, aquellas con mayor contribución para el CP1 fueron LIN (r = 0.84), EPR (r = 0.75) y LRM (r = 0.78). Este componente se caracterizó por agrupar variables relacionadas con potencial para producción de semilla. En el CP2 las variables con mayor contribución fueron RPI (r = 0.52) y LRM (r = 0.78), variables asociadas también con potencial para producción de semilla. Debido a esto, los genotipos con valores altos en el CP1 y CP2 son los que presentan mayor potencial para producir semilla.
A través del ACP se logró diferenciar y seleccionar los siguientes genotipos con características sobresalientes: genotipo 1 originario de Aguascalientes con ALT de 1900 msnm, TEMP de 18.8 ºC y PPma de 544 mm; genotipo 7 originario de Aguascalientes a 1750 msnm, 18.2 ºC y 380 mm; genotipo 19 recolectado en Querétaro a 2200 msnm, 16.5 ºC y 476 mm; genotipo 64 de Durango a 1800 msnm 18.0 ºC y 584 mm; genotipo 84 de Jalisco a 2250 msnm, 16.6 ºC y 542 mm; genotipo 508 de Chihuahua a 1570 msnm, 17.1 ºC y 380 mm y genotipo 512 de Chihuahua a 1170 msnm, 19.4 ºC y 278 mm (Figura 1). Los genotipos 7, 84 y 512, además de mostrar atributos deseables para producción de semilla, destacaron por su producción forrajera sobresaliente en un estudio anterior (Morales-Nieto et al., 2008). Debido a las características mencionadas, estos materiales son candidatos para su registro y liberación como futuras variedades.
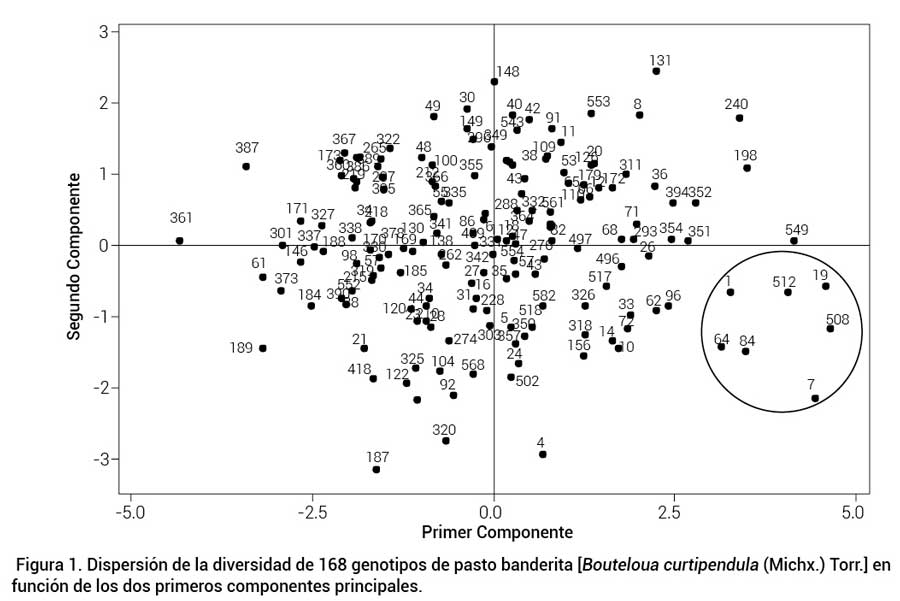
Figura 1.
Dispersión de la diversidad de 168 genotipos de pasto banderita [Boutelouacurtipendula (Michx.) Torr.] en función de los dos primeros componentes principales.
Análisis de Agrupamiento
La Figura 2 muestra el dendograma resultante del análisis de agrupamiento, donde los pseudo estadísticos F y T2 sugirieron cuatro grupos con un R2 = 0.76. Este coeficiente de determinación es considerado alto, tomando en cuenta el número de genotipos y la amplia variabilidad morfológica y productiva que mostraron. Los grupos I, II, III y IV quedaron conformados por 52, 32, 15 y 69 genotipos, respectivamente. En el grupo I se conjuntaron genotipos con alto potencial para producción de semilla. En el grupo II se integraron los genotipos con valores de medio a alto. El grupo III agrupó genotipos con valores de medio a bajo. Finalmente, el grupo IV agrupó los genotipos de menor potencial para producir semilla.
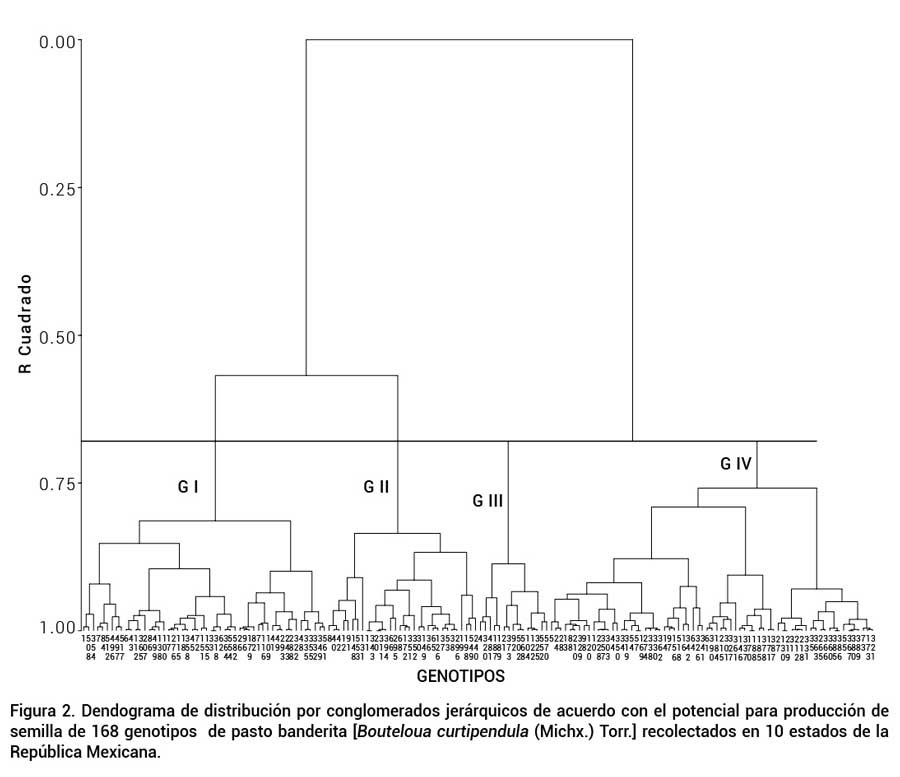
Figura 2
Dendograma de distribución por conglomerados jerárquicos de acuerdo con el potencial para producción de semilla de 168 genotipos de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recolectados en 10 estados de la República Mexicana.
Los análisis de CP y de agrupamiento coincidieron en integrar a los genotipos 1, 7, 84, 508 y 512 como los de mayor potencial para producción de semilla. De igual forma, Morales-Nieto et al. (2008) en un análisis de agrupamiento clasificaron estos genotipos en el grupo que presentó el mayor potencial para producción de forraje. No obstante, el grupo I presenta otros 47 genotipos que también pudieran tener aptitudes con potencial para producción de semilla.
Análisis de varianza multivariado
El análisis multivariante de la varianza (MANOVA) evidenció diferencias entre grupos (Lamda de Wilks = 0.099; P ≤ 0.0001), siendo todos los grupos diferentes ortogonalmente (P ≤ 0.0001). Es importante considerar que la capacidad productiva de una planta está relacionada con su expresión genética (Lobo, 2006). No obstante, la respuesta productiva depende en gran medida de las condiciones medioambientales y puede variar entre genotipos (Seymour et al., 2008; Tober et al., 2010). En este sentido, el utilizar estadísticos inferenciales da mayor certeza para afirmar que un genotipo o grupo de genotipos es diferente al resto de la población, independientemente de las condiciones ambientales de procedencia. Por lo anterior, se considera que el pasto banderita en México posee amplia variabilidad fenotípica, dentro de la cual se encuentran materiales con características relacionadas con producción de semilla.
Se han realizado estudios para caracterizar la diversidad morfológica en especies forrajeras, en los que se menciona que la caracterización morfológica de poblaciones naturales permite seleccionar genotipos adecuados para rehabilitar pastizales (Casler, 2005; Grenier et al., 2004; Morales et al., 2016); sin embargo, no se han realizado trabajos orientados a caracterizar la diversidad de atributos relacionados con producción de semilla.
Análisis de correspondencias
De acuerdo con el análisis de correspondencias, el gradiente de ALT del lugar de origen se asoció de manera significativa (P ≤ 0.1) con el agrupamiento de los genotipos. El grupo I presentó mayor correspondencia con los niveles bajo y medio de ALT. El grupo II se relacionó con altitudes bajas, mientras que el grupo IV se relacionó con ALT media. El grupo III se relacionó con altitudes mayores a los 2000 m (Figura 3).
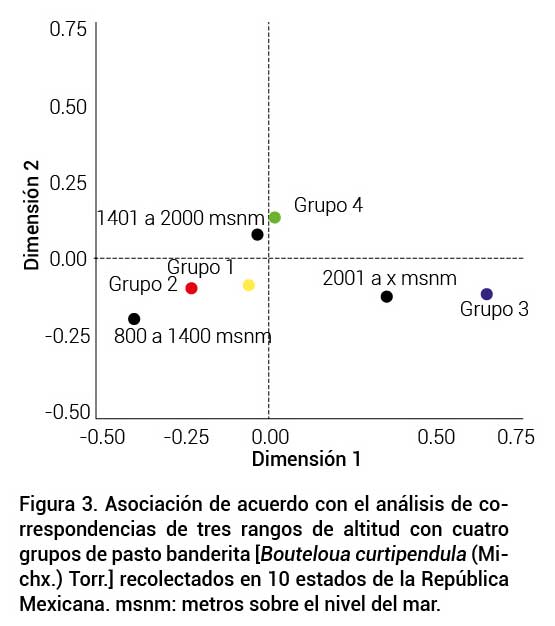
Figura 3
Asociación de acuerdo con el análisis de correspondencias de tres rangos de altitud con cuatro grupos de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recolectados en 10 estados de la República Mexicana. msnm: metros sobre el nivel del mar.
El análisis de correspondencia para el gradiente de PPma del lugar de procedencia de los genotipos mostró efecto significativo (P ≤ 0.05) sobre la conformación de los grupos. El grupo I presentó una alta correspondencia con el nivel de PPma alto. El grupo II se relacionó con el nivel de PPma bajo. El grupo III se relacionó con los niveles de PPma medio y alto, mientras que el grupo IV presentó correspondencia solamente con el nivel de PPma medio (Figura 4). Lo anterior concuerda con lo mencionado por Morales-Nieto et al. (2008), quienes señalaron que la diversidad morfológica entre genotipos puede deberse a diferencias en las condiciones ambientales del sitio de origen; sin embargo, a pesar de la relación entre las condiciones medioambientales de procedencia y la formación de los grupos, dentro de estos se presentaron genotipos recolectados de condiciones medioambientales diferentes. Tal es el caso del genotipo 512, el cual se ubicó dentro del grupo I; no obstante, es originario de un área de condiciones áridas. Esto probablemente se debe a que no todos los genotipos presentan correlación de sus características fenotípicas con las condiciones de origen (Jungmann et al., 2010; Vahabi et al., 2008). Otra explicación es que algunas gramíneas poseen alta plasticidad y se adaptan a diferentes condiciones medioambientales (Munson y Long, 2017).
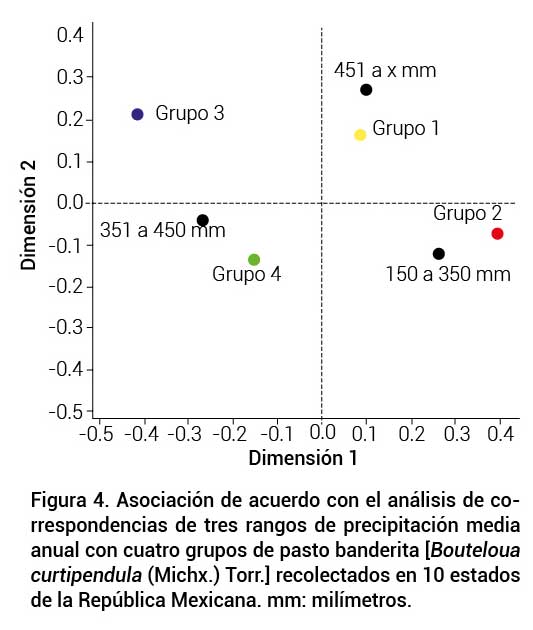
Figura 4.
Asociación de acuerdo con el análisis de correspondencias de tres rangos de precipitación media anual con cuatro grupos de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recolectados en 10 estados de la República Mexicana. mm: milímetros.
En la Figura 5 se observa cómo la distribución de grupos no mostró correspondencia con el origen geográfico de los genotipos (P > 0.05). Esto concuerda con lo señalado por diversos autores, quienes mencionan que, con base en las capacidades de expresión morfológica, es posible establecer grupos con características similares, más que por localidades o entidades de origen (Aguado-Santacruz et al., 2004; Avendaño et al., 2004; Núñez-Colín, 2010).
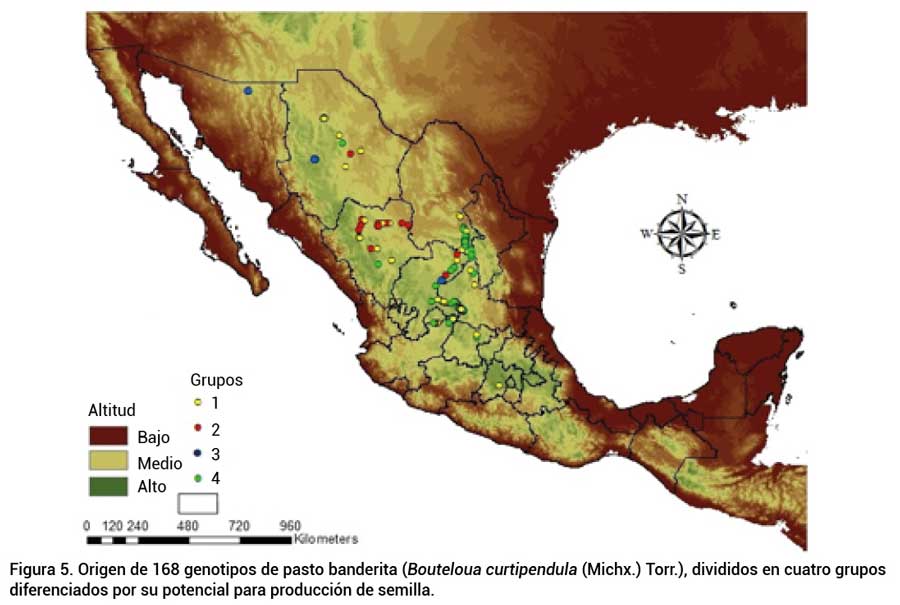
Figura 5
Origen de 168 genotipos de pasto banderita (Bouteloua curtipendula (Michx.) Torr.), divididos en cuatro grupos diferenciados por su potencial para producción de semilla
CONCLUSIONES
El complejo agámico Bouteloua curtipendula presentó variabilidad morfológica, lo cual permitió seleccionar genotipos con potencial para producción de semilla. Los genotipos 1, 7, 84, 508 y 512 fueron los más sobresalientes, por lo que estos materiales son candidatos para el registro de nuevas variedades y su uso en programas de restauración en zonas áridas y semiáridas de México.
Referencias
Aguado-Santacruz G. A., Q. Rascón C., J. L. Pons H., O. Grageda C. y E. García-Moya (2004) Manejo biotecnológico de gramíneas forrajeras. Técnica Pecuaria en México 42:261-276.
Avendaño A. C. H., P. Ramírez V., F. Castillo G., J. L. Chávez S. y G. Rincón E. (2004) Diversidad isoenzimática en poblaciones nativas de frijol negro. Revista Fitotecnia Mexicana 27:31-40.
Beltrán L. S., C. A. García D., J. A. Hernández A., C. Loredo O., J. Urrutia M., L. A. González E. y H. G. Gámez V. (2013) “Banderilla Diana” Bouteloua curtipendula (Michx.) Torr., nueva variedad de pasto para zonas áridas y semiáridas. Revista Mexicana de Ciencias Pecuarias 4:217-221.
Casler M. D. (2005) Ecotypic variation among switchgrass populations from the Northern USA. Crop Science 45:388-398.
Corrales L. R., C. R. Morales N., A. Melgoza C., J. S. Sierra T., J. Á. Ortega G. y G. Méndez Z. (2016) Caracterización de variedades de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] recomendadas para rehabilitación de pastizales. Revista Mexicana de Ciencias Pecuarias 7:201-211.
Ferdinandez Y. S. N. and B. E. Coulman (2004) Genetic relationships among smooth bromegrass cultivars of different ecotypes detected by AFLP markers. Crop Science 44:241-247.
Grenier C., P. J. Bramel, J. A. Dahlberg, A. E. Ahmadi, M. Mahmoud, G. C. Peterson, D. T. Rosenow and G. Ejeta (2004) Sorghums of the Sudan: analysis of regional diversity and distribution. Genetic Resources and Crop Evolution 51:489-500.
INEGI, Instituto Nacional de Estadística Geografía e Informática (2002) Estudio hidrológico del estado de San Luis Potosí. Instituto Nacional de Estadística Geografía e Informática. Aguascalientes, México. 124 p.
Jungmann L., B. B. Z. Vigna, K. R. Boldrini, A. C. B. Sousa, C. B. do Valle, R. M. S. Resende, M. S. Pagliarini, M. I. Zucchi and A. P. de Souza (2010) Genetic diversity and population structure analysis of the tropical pasture grass Brachiaria humidicola based on microsatellites, cytogenetics, morphological traits, and geographical origin. Genome 53:698-709.
Jurado-Guerra P., H. Domínguez-Caraveo, A. Melgoza-Castillo y C. R. Morales-Nieto (2012) Emergencia y crecimiento del zacate banderilla [Bouteloua curtipendula (Michx.) Torr.] con biosólidos en condiciones de sequía. Tecnociencia Chihuahua 6:57-67.
Lobo A. M. (2006) Recursos genéticos y mejoramiento de frutales andinos: una visión conceptual. Revista Corpoica-Ciencia y Tecnología Agropecuaria 7:40-54.
Martín R. M. H., F. A. Flores I., S. Moreno M., R. Retes L. y R. Amarillas R. (2016) Impacto económico asociado con la cosecha de semilla de zamota y mezquite en la región central de Sonora, México. Revista Mexicana de Agronegocios 38:217-228.
Morales N. C. R., C. H. Avendaño A., A. Melgoza C., K. C. Gil V., A. R. Quero C., P. Jurado G. y M. Martínez S. (2016) Caracterización morfológica y molecular de poblaciones de pasto banderita (Bouteloua curtipendula) en Chihuahua, México. Revista Mexicana de Ciencias Pecuarias 7:455-469.
Morales N. C. R. y A. Melgoza C. (2010) Características productivas de zacates forrajeros importantes en el norte de México. Folleto Técnico No. 28. S.E. La Campana-Madera. INIFAP-SAGARPA. Aldama, Chihuahua. 51 p.
Morales N. C. R., A. Melgoza C., P. Jurado G., M. Martínez S y C. H. Avendaño A. (2012) Caracterización fenotípica y molecular de poblaciones de zacate punta blanca [Digitaria californica (Benth.) Henr.]. Revista Mexicana de Ciencias Pecuarias 3:171-184.
Morales N. C. R., A. R. Quero C. y C. H. Avendaño A. (2007) Caracterización de la diversidad nativa del zacate banderita [Bouteloua curtipendula (Michx.) Torr.], mediante su nivel de ploidía. Técnica Pecuaria en México 45:263-278.
Morales N. C. R., L. Madrid P., A. Melgoza C., M. Martínez S., S. Arévalo G, Q. Rascón C. y P. Jurado G. (2009) Análisis morfológico de la diversidad del pasto [Bouteloua gracilis (Willd. ex Kunth) Lag. ex Steud.], en Chihuahua, México. Técnica Pecuaria en México 47: 245-256.
Morales-Nieto C. R., A. Quero-Carrillo, J. Pérez-Pérez, A. Hernández-Garay y O. Le-Blanc (2008) Caracterización morfológica de poblaciones nativas de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] en México. Agrociencia 42:767-775.
Morales-Nieto C. R., O. Rivero-Hernández, A. Melgoza-Castillo, P. Jurado-Guerra y M. Martínez-Salvador (2013) Caracterización morfológica y molecular de Leptochloa dubia (Poaceae) en Chihuahua, México. Polibotánica 36:79-94.
Munson S. M. and A. L. Long (2017) Climate drives shifts in grass reproductive phenology across the western USA. New Phytologist 213:1945-1955.
Núñez-Colín C. A. (2010) Distribución y caracterización eco-climática del membrillo cimarrón (Amelanchier denticulata (Kunth) Koch) en México. Revista Chapingo Serie Horticultura 16:195-206.
PACP-Ch, Plan de Acción para la Conservación de los Pastizales-Chihuahua (2011) Plan de Acción para la Conservación y Uso Sustentable de los Pastizales del Desierto Chihuahuense en el estado de Chihuahua 2011-2016. Gobierno del Estado de Chihuahua. Chihuahua, Chih. 44 p.
Quero-Carrillo A. R., F. J. Hernández-Guzmán, M. Velázquez-Martínez, H. G. Gámez-Vázquez, P. Landa-Salgado y P. Aguilar-López (2016) Métodos de establecimiento de pasturas en zonas áridas de México utilizando semillas crudas o cariópsides. Forrajes Tropicales 4:29-37.
Rowe C. L. J. and E. A. Leger (2012) Seed source affects establishment of Elymus multisetus in postfire revegetation in the Great Basin. Western North American Naturalist 72:543-553.
SAS Institute (2006) Statistical Analysis System 9.1.3 User´s guide. Cary, NC, USA. 210 p.
Schellenberg M. P., B. Biligetu, G. J. McLeod and Z. Wang (2012) Phenotypic variation of side-oats grama grass [Bouteloua curtipendula (Michx.) Torr.] collections from the Canadian prairie. Canadian Journal of Plant Science 92:1043-1048.
Seymour R., J. Seymour and C. Blackford (2008) Six basic elements for a successful native grass and forb establishment. 3rd ed. Roundstone native seed, LLC. Upton, Kentucky. USA. 24 p.
Tober D., N. Jensen, W. Duckwitz and M. Knudson (2010) Sideoats grama (Bouteloua curtipendula). Performance trials North Dakota, South Dakota, and Minnesota. USDA, Natural Resources Conservation Service, Bismarck, North Dakota. USA. 16 p.
Vahabi A. A., A. Lotfi, M. Solouki and S. Bahrami (2008) Molecular and morphological markers for the evaluation of diversity between Plantago ovata in Iran. Biotechnology 7:702-709.
Whalley R. D. B., I. H. Chivers and C. M. Waters (2013) Revegetation with Australian native grasses-a reassessment of the importance of using local provenances. The Rangeland Journal 35:155-166.
Notas de autor
esantellanoe@gmail.com