ARTÍCULOS CIENTÍFICOS
DIVERSIDAD GENÉTICA DE CHILE HABANERO (Capsicum chinense Jacq.) MEDIANTE ISSR
GENETIC DIVERSITY OF HABANERO PEPPER (Capsicum chinense Jacq.) USING ISSR
DIVERSIDAD GENÉTICA DE CHILE HABANERO (Capsicum chinense Jacq.) MEDIANTE ISSR
Revista Fitotecnia Mexicana, vol. 41, núm. 3, pp. 227-236, 2018
Sociedad Mexicana de Fitogenética, A.C.
Recepción: 10 Abril 2017
Aprobación: 23 Mayo 2018
Resumen: El chile habanero (Capsicum chinense Jacq.) es distintivo de la Península de Yucatán, México; sin embargo, los estudios de diversidad genética a nivel molecular en este cultivo son escasos. El presente trabajo se planteó con el objetivo de caracterizar la diversidad genética del germoplasma de chile habanero, mediante marcadores ISSR. El material genético consistió en 60 poblaciones criollas de chile habanero recolectadas en los estados de Yucatán, Campeche, Quintana Roo y Tabasco, México, las cuales fueron genéticamente analizadas con tres loci ISSRs. El ADN fue extraído por el método de CTAB y los fragmentos se amplificaron por la reacción en cadena de la polimerasa (PCR). Los datos se analizaron con los programas POPGENE V 1.31 y NTSYS V 2.0 mediante un análisis de componentes principales y las relaciones entre las poblaciones se obtuvieron con un análisis jerárquico de conglomerados usando el método UPGMA. Se detectaron un total 32 bandas y 98 % de ellas fueron polimórficas. Los resultados indican que la diversidad genética en poblaciones de chile habanero es alta, en donde el 95.5 % de la variación observada se encuentra dentro de las poblaciones y sólo el 4.5 % entre ellas. Se detectó un alto flujo génico entre las poblaciones. Los resultados de los análisis multivariados sugieren que gran cantidad de germoplasma de chile habanero se ha dispersado del estado de Yucatán a los estados de Campeche, Quintana Roo y Tabasco. El patrón de distribución de la diversidad genética de chile habanero no se asoció con el lugar de origen de las poblaciones analizadas.
Palabras clave: Capsicum chinense, diversidad genética, caracterización molecular, marcadores ISSR.
Abstract: Habanero pepper (Capsicum chinense Jacq.) is distinctive of the Yucatan Peninsula, Mexico; however, studies of genetic diversity at the molecular level on this crop are scarce. The present study was proposed with the objective of characterizeing the genetic diversity of habanero pepper germplasm by using ISSR markers. The genetic material consisted of 60 landrace populations of habanero pepper collected in the states of Yucatan, Campeche, Quintana Roo and Tabasco, Mexico, which were genetically analyzed with three ISSR loci. DNA was extracted by the CTAB method, and the fragments were amplified through the polymerase chain reaction (PCR). Data were analyzed using the POPGENE V 1.31 and NTSYS V 2.0 programs by means of principal component analysis, and the genetic relationships among populations were obtained by hierarchical cluster analysis based on the UPGMA method. A total of 32 bands were detected and 98 % of them were polymorphic. Results indicate that genetic diversity in habanero pepper is large, 95.5 % of the observed variation occurred within the populations and only 4.5 % among them. A high gene flow was detected among populations. Results of multivariate analyses suggest that a large amount habanero pepper germplasm has dispersed from the state of Yucatan to the states of Campeche, Quintana Roo and Tabasco. The distribution pattern of genetic diversity of habanero pepper populations was not associated to the provenance of the analyzed populations.
Keywords: Capsicum chinense, genetic diversity, molecular characterization, ISSR markers.
INTRODUCCIÓN
El género Capsicum lo integran alrededor de 35 especies, dentro de las cuales las de mayor importancia por su domesticación y superficie cultivada son Capsicum annuum L., Capsicum frutescens L., Capsicum pubescens Ruíz & Pavón, Capsicum baccatum L. y Capsicum chinense Jacq. (Bosland y Votava, 2012; Carrizo et al., 2013). México es centro de origen y diversidad de C. annuum y además se encuentran presentes las especies C. frutescens, C. pubescens y C. chinense. En este sentido Aguilar et al. (2010) reportan para México 56 tipos de chile entre cultivados, semicultivados y silvestres. Recientemente Vera-Sánchez et al. (2016) mencionan 64 tipos de chile (95 % de éstos pertenecen a C. annuum), los que se distribuyen principalmente en las zonas centro, sur y sureste de México.
En México el chile es el octavo cultivo en cuanto a valor económico, con un volumen promedio de producción de 2.2 millones de toneladas anuales, de las cuales se exportan cerca de 900 mil en diferentes presentaciones (fresco, seco, polvo, entre otros) (SIAP, 2018). La importancia de este cultivo radica en su valor económico, alimenticio (especias y vegetales) y como generador de mano de obra. Es materia prima para la industria alimentaria, farmacéutica, cosmética e industrial (Aza-González et al., 2011).
En particular, el chile habanero (C. chinense Jacq.) es distintivo de la Península de Yucatán, México. Fue introducido por el Caribe, se adaptó a las condiciones agroecológicas y al manejo tradicional de los agricultores yucatecos; además, se integró a la cultura y gastronomía de la región, de donde se dispersó a todo el país, y actualmente se siembra a nivel comercial en, al menos, 17 estados (Latournerie et al., 2002). El chile habanero tiene características propias como el aroma, sabor, consistencia, picor, entre otras; con base en lo anterior, el 4 de junio de 2010 se otorgó la denominación de origen como “Chile habanero de la Península de Yucatán” (IMPI, 2010) y posteriormente se originó la Norma Oficial Mexicana PROY-NOM-189-SCFI-2012 Chile habanero de la Península de Yucatán (Capsicum chinense Jacq.) Especificaciones y métodos de prueba (Secretaría de Economía, 2012). En la Península de Yucatán se producen alrededor de 3500 t anuales que son comercializadas principalmente en fresco y en menor grado procesadas.
La variabilidad del chile habanero ha sido analizada vía caracterización fenotípica (Latournerie et al., 2002), caracterización isoenzimática (Corona-Torres et al., 2000; Howard et al., 2000), evaluación agronómica (Latournerie-Moreno et al., 2015) y caracterización bioquímica (Coutinho et al., 2015; Guzman et al., 2010). Los trabajos a nivel molecular son escasos; por ejemplo, se han realizado estudios de diversidad entre las cinco especies de Capsicum, donde se incluye al chile habanero como un componente (Dias et al., 2013; Lijun y Xuexiao, 2012); pero no se tienen reportes de estudios de la diversidad genética existente exclusivamente en el germoplasma de chile habanero.
Como complemento a la caracterización morfológica se utilizan los marcadores moleculares. Entre los diferentes tipos de marcadores para estudiar la diversidad genética sobresalen los SSR (Simple Sequence Repeats) y los ISSR. Los SSR son codominantes y se les considera ideales para la identificación genética individualizada (Jones et al., 2010); además, tienen cobertura extensiva del genoma, localización específica en los cromosomas, entre otras ventajas. Una de las desventajas de estos marcadores es que requiere considerable infraestructura en laboratorio y presupuesto (Kalia et al., 2011). Los ISSR, son segmentos de ADN localizados entre secuencias repetitivas de nucleótidos, que generalmente aparecen en regiones no codificantes del genoma, son dominantes, fácilmente reproducibles y no afectados por el ambiente (Laurentin, 2009), y son de bajo costo para evaluar diversidad genética (Zizumbo-Villareal et al., 2005); además, permiten realizar estudios intra e interespecíficos. El objetivo de este trabajo consistió en caracterizar la diversidad genética de germoplasma de chile habanero de la Península de Yucatán y Tabasco, México, mediante marcadores genéticos del tipo ISSR.
MATERIALES Y MÉTODOS
Material vegetal
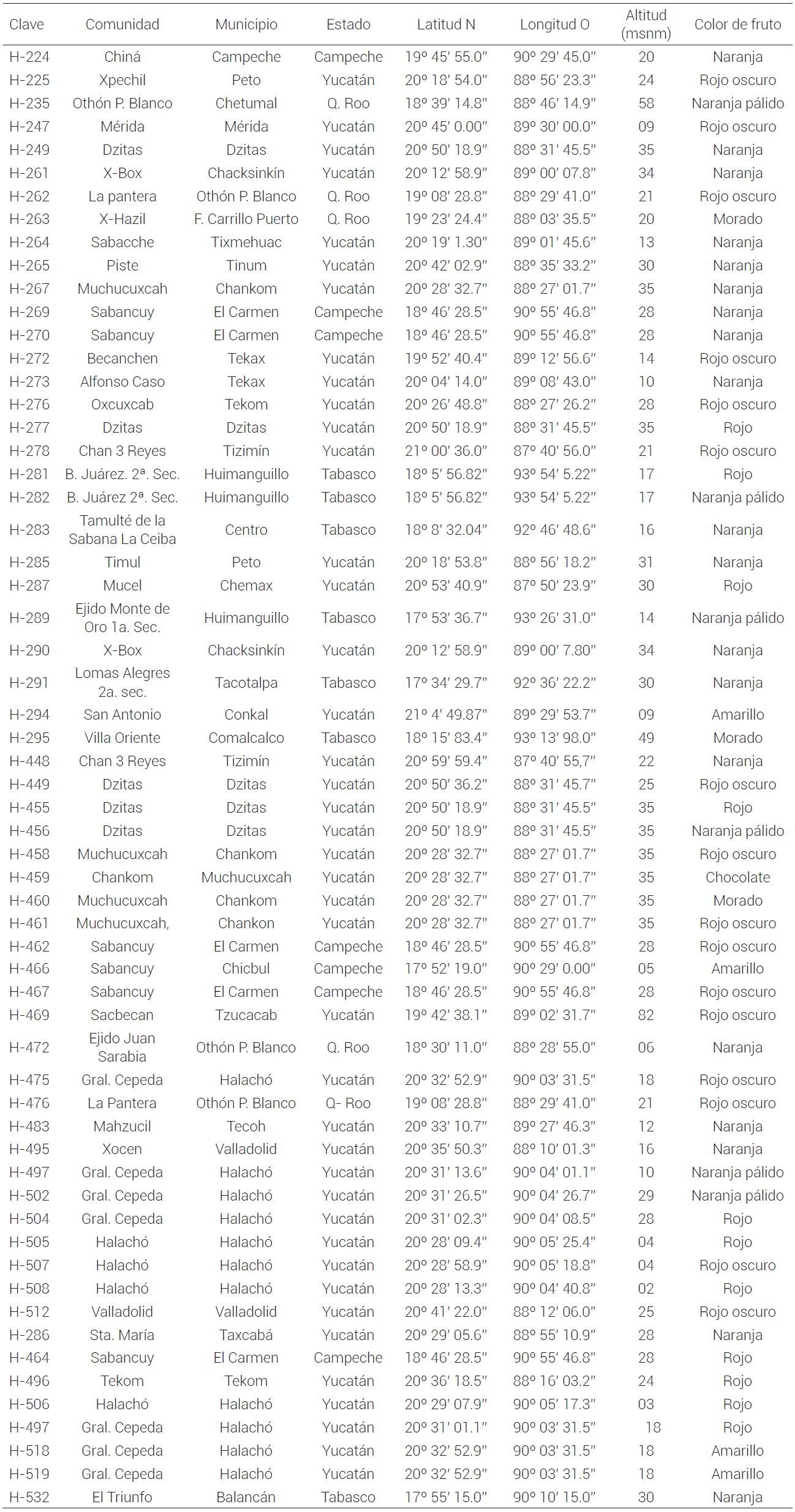
El germoplasma estuvo conformado por 60 accesiones de chile habanero que se recolectaron en los estados de Yucatán (40), Campeche (7), Quintana Roo (5) y Tabasco (8). Con base en el color de los frutos a la madurez, 27 fueron de color rojo, 25 naranja, 4 amarillo y 4 morado (Cuadro 1). Las accesiones provienen de los trabajos de recolección realizados por la Red de Chile del SINAREFI-SNICS-SAGARPA (Sistema Nacional de Recursos Fitogenéticos para la Alimentación y la Agricultura-Servicio Nacional de Inspección y Certificación de Semillas-Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación) entre 2006 y 2014. La estrategia de colecta se basó en el color de frutos a la madurez y se recolectó un máximo de cinco accesiones por comunidad por variante identificada, así como por la región donde se siembran. Cada accesión se constituyó de al menos 20 plantas muestreadas al azar en los campos de cultivo y en el caso de solares o traspatio de cinco a 10 plantas. Para el presente trabajo cada colecta o accesión se consideró como una población, dado que es representativa de un área determinada con base en la estrategia de recolección usada.
Extracción de ADN
Se utilizaron tres hojas de cinco plantas de cada accesión (1 g de muestra) de una edad de 45 días después de la siembra (sembradas en condiciones protegidas el 15 de junio de 2015). Para la extracción de ADN genómico se utilizó el protocolo de bromuro de cetil trimetil amonio (CTAB) descrito por Doyle y Doyle (1987), modificado al 3 % para C. chinense. La integridad del ADN fue verificada por medio de electroforesis en gel de agarosa al 0.8 %, la concentración mediante absorbancia a 260 nm y la pureza a través de la relación de absorbancias 260/280.
Amplificación de ADN por PCR
Se evaluaron inicialmente 17 iniciadores ISSR (InvitrogeneTM), de éstos solamente se seleccionaron los tres que amplificaron de manera consistente, clara y que presentaron el mayor nivel de polimorfismo. Cada banda ISSR fue considerada como un locus independiente (Cuadro 2). La amplificación de los fragmentos de ADN se realizó en un termociclador (GeneAmp® PCR System 9700, Foster City, CA, USA). Las reacciones de PCR fueron realizadas en un volumen final de 25 µL, que contenían 20 pmol de iniciador, buffer de reacción (20 mM Tris-HCl, pH 8, 50 mM KCl), 2U de Taq ADN polimerasa, 2.0 mM MgCl2, 10 mM dNTP y 40 ng de ADN genómico. Los perfiles de temperatura fueron de 5 min a 94 ºC, seguidos de 35 ciclos
de: 1 min a 94 ºC, 1 min a 53, 49 o 44 ºC (dependiendo del iniciador) y 2 min a 72 ºC, con extensión final de 5 min a 72 ºC. Los productos amplificados fueron visualizados en gel de agarosa al 2 % con amortiguador TAE 1X, pH 8, teñidos con bromuro de etidio y visualizados en un transiluminador UV (Benchtop UV trasluminator UVP®, Upland, CA, USA).
Análisis de datos genéticos
Los datos se registraron en una matriz binaria 1 y 0; donde el 1 indicó presencia y el 0 ausencia de bandas. El análisis de la información se realizó con el programa POPGENE V 1.31 (Yeh et al., 1999); se calcularon los siguientes parámetros: 1) diversidad genética mediante el porcentaje de loci polimórficos (PLP), 2) la diversidad génica de Nei (He), índice de diversidad de Shannon (I), 3) diversidad intrapoblacional (Hs), 4) flujo genético entre poblaciones (Nm) y 5)

R = (A, G); Y = (C, T).
índice de diferenciación entre poblaciones (Gst). Se realizó un análisis de componentes principales (ACP) y un análisis de conglomerados jerárquicos por el método UPGMA (Unweighted Pair Group Method with Arithmetic Averages), calculado con las distancias Euclidianas como medida de similitud, con el programa NTSYS (Numerical Taxonomic and Multivariate Analysis System) version 2.0 (Rohlf, 1998).
RESULTADOS Y DISCUSIÓN
Variabilidad genética
El análisis de 60 poblaciones de chile habanero con tres iniciadores ISSR seleccionados permitió identificar un total de 32 bandas, cada una considerada como un locus; el tamaño de las bandas varió de 300 a 1500 pb (Figura 1) y 98 % de loci polimórficos (Cuadro 3). Al respecto, en México se han realizado varios estudios relacionados con diversidad genética en diferentes tipos de chiles cultivados y silvestres de C. annuum; entre ellos, Contreras et al. (2011) en 55 poblaciones de chile Poblano del estado de Puebla reportaron 105 alelos en total y 5.53 alelos por locus para 19 loci SSR y polimorfismo de 80 %, el cual es inferior al encontrado en el presente trabajo. Toledo-Aguilar et al. (2016) utilizaron 24 loci de SSR, para analizar la diversidad genética de chile ancho y encontraron 220 alelos, con un promedio de 9.2 alelos por locus en 44 accesiones de ese tipo de chile.
Con el mismo tipo de marcador SSR, también se han reportado estudios de diversidad genética entre diferentes especies de Capsicum; por ejemplo, Dhaliwal et al. (2014), en 64 accesiones de tres especies de Capsicum de la India (C. annuum, C. Chinense y C. frutescens) utilizaron 27 loci SSR y obtuvieron 75 alelos en total y 2.78 alelos por locus. Por su parte, Hanáček et al. (2009) analizaron 41 accesiones de diferentes tipos de C. annuum de la India con 8 loci SSR y reportaron 28 alelos en total, 3.5 alelos por locus, con 62.5 % de loci polimórficos. Ibiza et al. (2012) observaron un promedio de 14.5 alelos por locus con 10 loci SSR en 270 poblaciones de la región de los Andes pertenecientes a cinco especies de Capsicum (C. annuum, C. chinense, C. frutescens, C. baccatum y C. pubescens). En general, los diferentes estudios sugieren que el uso de un mayor número de iniciadores no implica encontrar un mayor número de alelos por locus. Lo anterior refuerza la importancia de usar marcadores moleculares específicos que permitan identificar mayor polimorfismo dentro y entre las poblaciones bajo estudio.
La heterocigosidad esperada (He) en poblaciones de chile habanero fue de 0.40. Al respecto, diversos estudios reportan mayor He con microsatélites (SSR) en poblaciones silvestres de chile (C. annuum var. grabiusculum), cuyos valores de He (0.47 a 0.78) superan los de chiles cultivados (C. annuum L.) que varían entre 0.42 y 0.59 (Hernández, 2014; Nicolaï et al. 2013; Pacheco-Olvera et al., 2012).
El índice de diversidad de Shannon (I) en el germoplasma de chile habanero analizado fue de 0.59 (Cuadro 3), el cual
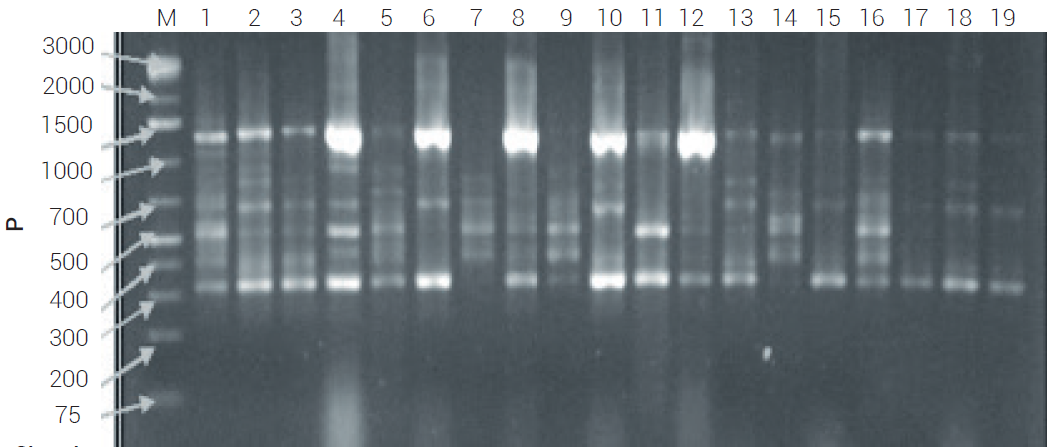
Figura 1
Perfiles de ISSR detectados con el iniciador (AC)8YG en 19 poblaciones de chile habanero (Capsicum chinense Jacq.) en gel de agarosa al 2 %. El carril M corresponde a la escalera molecular.
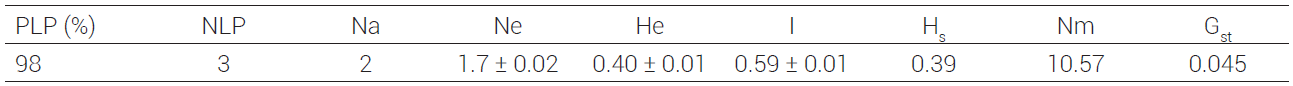
PLP: porcentaje de loci polimórficos, NLP: número de loci polimórficos, Na: número de alelos observados, Ne: número de alelos esperados, He: heterocigosidad esperada (Índice de Nei, 1973), I: índice de diversidad de Shannon, Hs: heterocigosidad intrapoblacional, Nm: flujo génico, Gst: coeficiente de diferenciación genética.
indica una diversidad intermedia entre las poblaciones de chile habanero estudiadas. Los resultados del presente estudio superan el valor de 0.33 reportado por Lijun y Xuexiao (2012), en el cual incluyeron las cinco especies domesticadas de Capsicum (Hanáček et al., 2009). Al respecto, Dawson et al. (1995) indican que el índice de diversidad de Shannon no es sensible a efectos del sesgo causado por la incapacidad de detectar a heterocigotos.
Diferenciación genética en las poblaciones
Los valores del índice de diferenciación genética (Gst) indican la proporción de la diversidad genética entre las poblaciones involucradas (Sounigo et al., 2005). La diferenciación genética entre las poblaciones de chile habanero de la Península de Yucatán y Tabasco resultó baja (Gst = 0.045), lo que indica que las poblaciones están poco diferenciadas, y que la mayor proporción de la diversidad genética (95.5 %) está representada dentro de las poblaciones de chile habanero (variación intra-poblacional) (Cuadro 3). Eguiarte et al. (2015) mencionan que, desde el punto de vista molecular, se esperarían bajos valores de diversidad genética de una pequeña muestra de la población que proviene de una reducida distribución geográfica de un material endémico. Lijun y Xuexiao (2012) observaron valores de Gst = 0.17 en cinco especies de Capsicum, mientras que Oyama et al. (2006) reportaron un valor de Gst de 0.32 en C. annuum (chile Serrano, Jalapeño y Morrón) del noroeste de México, el cual supera por mucho al encontrado en esta investigación con C. chinense.
De los resultados mencionados, se puede establecer que hay mayor diferenciación entre poblaciones que pertenecen a diferentes tipos de chile que entre poblaciones que corresponden a un mismo tipo de chile y de una región en particular, como es el caso del chile habanero. Al respecto, Lijun y Xuexiao (2012) y Oyama et al. (2006) mencionan que estos resultados están influenciados por el tipo de marcador utilizado, número de muestras analizadas, origen de las muestras, y por los diferentes tipos de chiles analizados y reportados en la literatura. En las poblaciones estudiadas de chiles habanero se encontró baja heterocigosidad intrapoblacional (Hs = 0.39) lo que indica moderada variabilidad genética en las poblaciones.
La baja diferenciación genética observada entre poblaciones de chile habanero se explica por el elevado flujo génico (Nm = 10.57) detectado entre las 60 poblaciones analizadas. Lo anterior indica que existen al menos 10 individuos migrantes en los materiales analizados. Estos resultados se explican en parte por las altas tasas de polinización cruzada que se reportan en C. chinense (Corona-Torres et al., 2000), lo cual depende en gran medida de la biología floral, la temperatura y la presencia de insectos polinizadores (abejas principalmente), durante la etapa de floración. También contribuyen a la baja diferenciación genética otros factores como la anatomía de la planta, barreras geográficas, expresión genética e interacción genotipo-ambiente (Eguiarte et al., 1993).
Relación genética interpoblacional
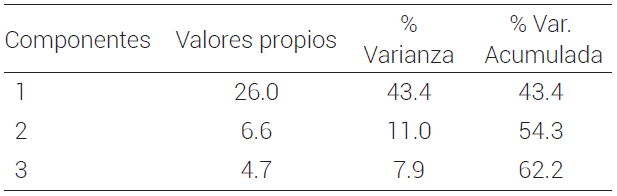
Con base en el análisis de componentes principales (ACP), los tres primeros componentes explicaron el 62.24 % de la variación genética total observada. El CP1 contribuyó con el 43.4 % de la variación total, mientras que los CP2 y CP3 aportaron el 11 y 7.9 % de la variación total acumulada, respectivamente (Cuadro 4). La variación observada en los primeros tres componentes concuerda con la baja estructura genética encontrada en las poblaciones estudiadas. Valores inferiores fueron reportados por Contreras et al. (2011) en chile poblano y ancho (C. annum L.), así como Toledo-Aguilar et al. (2016) con chile ancho (C. annum L.). Sus valores explicaron el 50.5 % de la variación total observada, con 10 componentes principales, para lo cual usaron marcadores microsatélites (SSR).
Con base en un plano cartesiano generado por los componentes principales CP1 y CP2 (Figura 2) el germoplasma evaluado se integró en tres grupos (I, II, III) y siete poblaciones (H262, H224, H504, H448, H458, H497 y H449) quedaron en forma independiente. El grupo I, integrado por 29 poblaciones, de las cuales el 65 % son originarias del estado de Yucatán, 13 % de Tabasco, 10 % de Campeche y 6 % de Quintana Roo. El grupo II se constituyó por ocho poblaciones, de las cuales seis (75 %) son originarias del estado de Yucatán, una (12.5 %) de Tabasco y una (12.5 %) de Campeche. El grupo III quedó conformado por 16 poblaciones, la mayoría de estas provenientes de Yucatán (62 %) y el resto de Tabasco (16 %), Quintana Roo (16 %) y Campeche (16 %).
Con base en el ACP de las 60 poblaciones evaluadas se puede establecer que el germoplasma de chile habanero ha migrado de Yucatán a las otras áreas de colecta. Esta migración es posible que se haya dado por semilla y por plántulas transportadas de Yucatán a Quintana Roo y Campeche principalmente, por lo que estas poblaciones están genéticamente más relacionadas dentro de grupos. Resultados similares fueron reportados por McLeod et al. (1982) con isoenzimas, quienes sugirieron que una mayor proporción del género Capsicum surgió de la parte central del sur de Bolivia, con una subsecuente migración hacia los Andes y a las tierras bajas del Amazonas.
Análisis de similitud
Los resultados del análisis de agrupamiento por el
método de UPGMA con las 60 poblaciones de chile habanero (Figura 3) se complementan con el ACP. El método UPGMA mostró tres grupos bien definidos (Grupos 2, 3 y 4) y tres grupos independientes (Grupos 1, 5 y 6). El grupo 2 se integró por tres subgrupos bien diferenciados, donde sobresale el subgrupo integrado por dos poblaciones (H247 y H281) originario del estado de Yucatán, y que se separan del resto del grupo 2. También se observó que dentro del grupo 2, las poblaciones de Yucatán H249 y H261 son idénticas genéticamente entre sí. Esto puede estar relacionado con el sistema informal de semillas que usan los agricultores para satisfacer sus necesidades para la siembra, el cual constituye un sistema dinámico muy importante para mantener la diversidad que cultivan los agricultores de subsistencia (Poudel et al., 2015). El movimiento de semilla se da principalmente entre familiares, conocidos, etc., por lo que una variedad puede ser desplazada de una región a otra (Hodgkin et al., 2007). Sobresale el grupo 4, por ser el único grupo que se integró con poblaciones de Yucatán (H495, H502, H504 y H512) y que se separa del resto de los grupos, lo que indica mayor diferenciación genética de este grupo.
Las poblaciones H224 originaria del estado de Campeche (grupo 1), H262 originaria del estado de Quintana Roo (grupo 5) y H458 del estado Yucatán (grupo 6) se agruparon de forma independiente, las cuales presentaron la mayor diferenciación genética entre el germoplasma estudiado del chile habanero, lo que es indicativo que la diversidad se está generando entre los estados de la Península de Yucatán y en particular en mayor grado dentro del estado de Yucatán; es decir, el flujo de genes entre estas poblaciones es limitado debido al aislamiento geográfico de las áreas donde se cultiva cada población. Resultados similares fueron reportados por Naino Jika et al. (2017) quienes encontraron un reducido flujo de genes entre variedades criollas manejadas por diferentes grupos étnicos en África.
Con base en el ACP y análisis de agrupamiento con ISSR, se puede señalar que la diversidad en chile habanero en general no se asocia con el lugar de origen de las poblaciones (Cuadro 1), lo cual coincide con lo obtenido por Lijun y Xuexiao (2012), quienes estudiaron la variabilidad genética en cinco especies de Capsicum con 15 iniciadores ISSR (entre ellos AC8YG, utilizado en el presente estudio), donde concluyeron que el agrupamiento jerárquico no se asoció con el sitio de origen de las variedades estudiadas. De la misma forma, no se encontró relación entre el color del fruto y la diversidad observada a nivel de marcadores moleculares. Por otro lado, la mayoría de las poblaciones de Campeche y Quintana Roo, y todas las de Tabasco (H281,
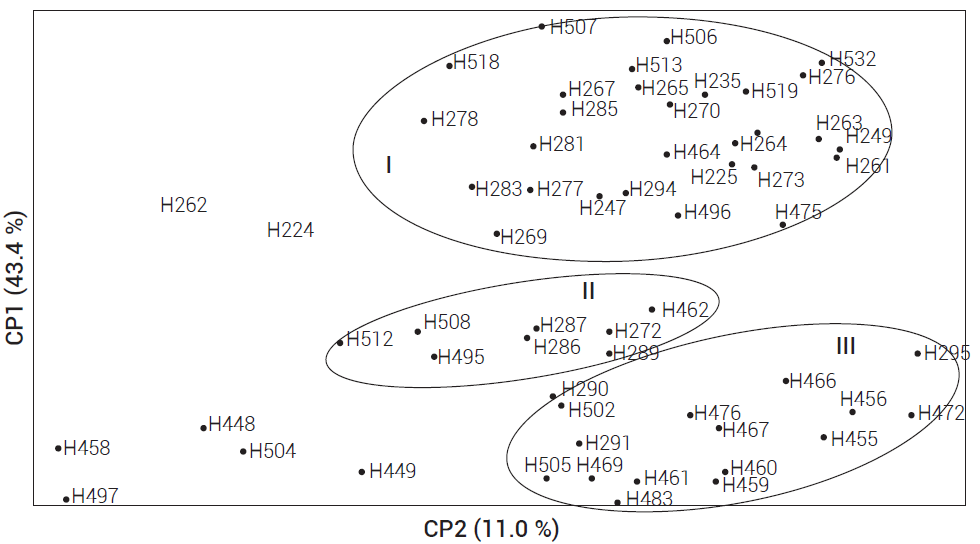
Figura 2
Dispersión de 60 poblaciones de chile habanero con base en los dos primeros componentes principales (CP1 y CP2) obtenidos de 32 alelos con tres iniciadores ISSR.
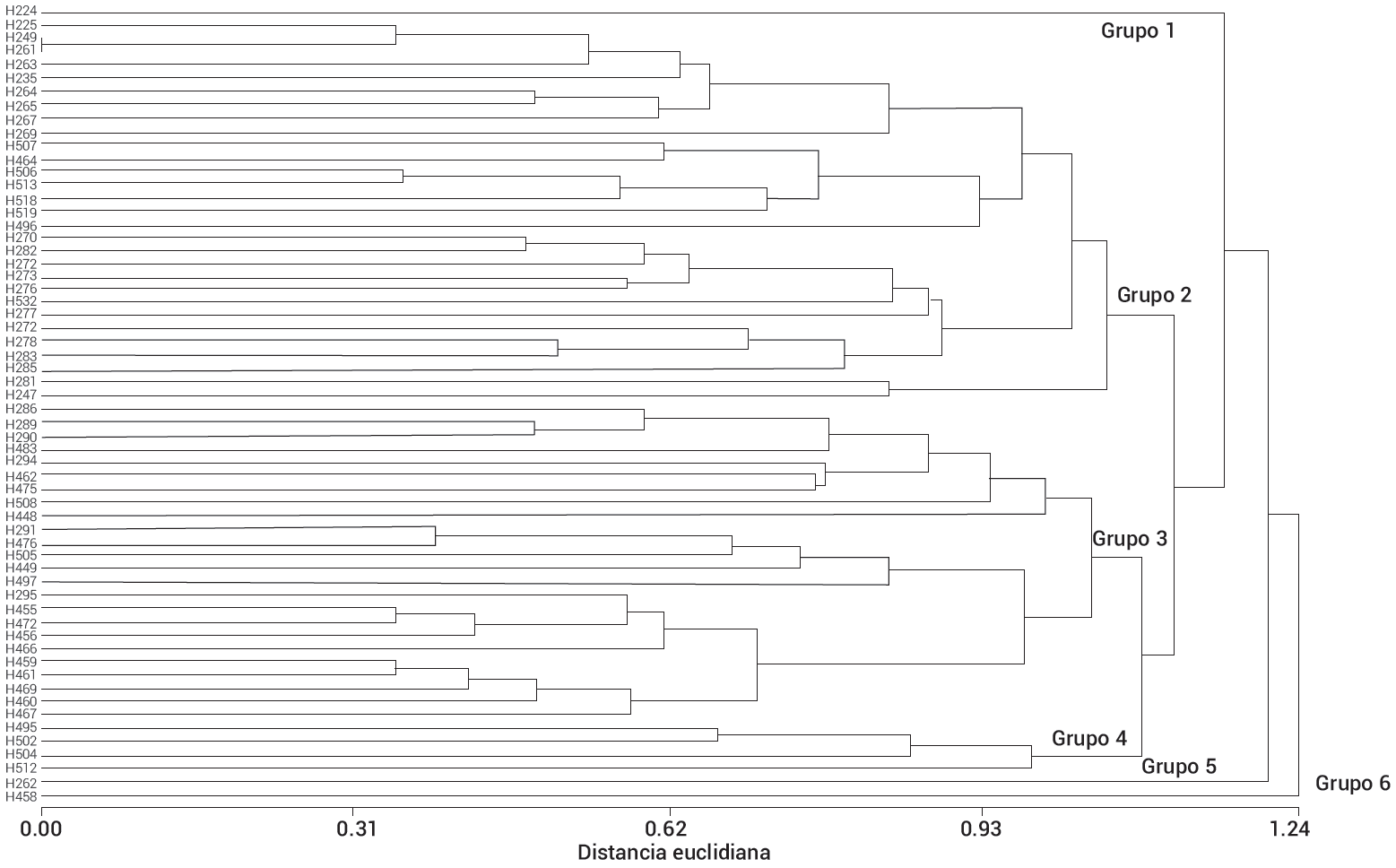
Figura 3
Dendrograma jerárquico con el método UPGMA que representa el agrupamiento genético de 60 poblaciones de chile habanero recolectadas en la Península de Yucatán y Tabasco.
H282, H283, H289, H291, H295 y H532) quedaron integradas genéticamente con las poblaciones de Yucatán, lo cual sugiere que se originaron a partir de éstas. Las diferentes condiciones ambientales y de manejo han generado ciertos cambios fenotípicos entre las poblaciones que salieron de Yucatán, pero no se detecta una fuerte diferenciación genética entre ellas.
CONCLUSIONES
La diversidad genética en las poblaciones de chile habanero colectadas en la Península de Yucatán y Tabasco es relativamente alta. El germoplasma de chile habanero muestra mayor variación genética dentro de las poblaciones que entre ellas y se presenta alto flujo génico entre las poblaciones. Las poblaciones de chile habanero del estado de Yucatán presentan la mayor variación genética, lo cual confirma al estado de Yucatán como centro de dispersión de chile habanero en el sureste mexicano. No se encontró relación entre la diversidad genética observada y el lugar de origen de las poblaciones.
Referencias
Aguilar R. V. H., T. Corona T., P. López L., L. Latournerie M., M. Ramírez M., H. Villalón M. y J. A. Aguilar C. (2010) Los chiles de México y su Distribución. SINAREFI, Colegio de Postgraduados, INIFAP, IT-Conkal, UANL, UAN. Montecillo, Texcoco, Estado de México. 114 p.
Aza-González C., H. G. Núñez-Palenius and N. Ochoa-Alejo (2011) Molecular biology of capsaicinoid biosynthesis in chili pepper (Capsicum spp.). Plant Cell Reports 30:695-706. http://doi.org/10.1007/s00299-010-0968-8
Bosland P. W. and E. y J. Votava (2012) Peppers: Vegetable and Spice Capsicums. Crop Production Science in Horticulture. 2nd edition. CABI Publishing. Wallinford, UK. 230 p.
Carrizo G. C., M. Sterpetti, P. Volpi, M. Ummarino and F. Saccardo (2013) Wild capsicums: identification and in situ analysis of Brazilian species. In: Breakthroughs in the Genetics and Breeding of Capsicum and Eggplant. Proceedings of the XV Eucarpia Meeting on Genetics and Breeding of Capsicum and Eggplant, Torino, Italy. 2-4 September 2013. S. Lanteri and G. L. Rotino (eds.). I Love Books. Delmar, New York. pp:205-213.
Contreras T. A. R., H. López S., A. Santacruz V., E. Valadez M., V. H. Aguilar R., T. Corona T. y P. A. López (2011) Diversidad genética en México de variedades nativas de chile ‘Poblano’ mediante microsatélites. Revista Fitotecnia Mexicana 34:225-232.
Corona-Torres T., A. García-Velázquez, F. Castillo-González, V. Montero-Tavera y H. S. Azpiroz-Rivero (2000) Caracterización isoenzimática de la diversidad genética de colectas de chile (Capsicum annuum L. y Capsicum chinense Jacq.). Revista Chapingo Serie Horticultura 6:5-17, http//dx.doi.org/10.5154/r.rchsh.1999.04.033
Coutinho J. P., G. F. Barbero, O. F. Avellán, A. Garcés- Claver, H. T. Godoy, M. Palma and C. G. Barroso (2015) Use of multivariate statistical techniques to optimize the separation of 17 capsinoids by ultra performance liquid chromatography using different columns. Talanta 134:256–263. https://doi.org/10.1016/j.talan-ta.2014.11.004
Dawson I. K., A. J. Simons, R. Waugh and W. Powell (1995) Diversity and ge- netic differentiation among subpopulations of Gliricidia sepium revealed by PCR-based assays. Heredity 74:10-18, https//doi.org/10.1038/hdy.1995.2
Dhaliwal M. S., A. Yadav and S. K. Jindal (2014) Molecular characterization and diversity analysis in chilli pepper using simple sequence repeats (SSR) markers. African Journal of Biotechnology 13:3137-3143, https://doi.org/10.5897/AJB2014.13695
Dias G. B., V. M. Gomes, T. M. S Morales, U. P. Zottich, G. R. Rabelo, A. O. Carvalho, M. Moulin, L. S. A. Gonçalves, R. Rodrigues and M. Da Cunha (2013) Characterization of Capsicum species using anatomical and molecular data. Genetics and Molecular Research 12:6488- 6501. https://doi.org/10.4238/2013.February.28.29
Doyle J. J. and J. L. Doyle (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissues. Phytochemical Bulletin 19:11-15.
Eguiarte L. E., A. Búrquez, J. Rodríguez, M. Martínez-Ramos, J. Sarukhán and D. Piñero (1993) Direct and indirect estimates of neighborhood and effective population size in a tropical palm, Astrocaryum mexicanum. Evolution 47:75-87, https://doi.org/10.1111/j.1558-5646.1993.tb01200.x
Eguiarte L. E., E. Aguirre-Planter, E. Scheinvar, A. González G. and V. Sousa (2015) Diferenciación, flujo génico y estructura genética de las poblaciones con énfasis en especies de plantas mexicanas. In: Evolución Orgánica. A. Becerra, A. Castañeda y D. Piñero (coords.). Universidad Nacional Autónoma de México. México, D. F. pp:10-13.
Guzman I., S. Hamby, J. Romero, P. W. Bosland, and M. A. O’Connell (2010) Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Science 179:49-59, https://doi.org/10.1016/j.plantsci.2010.04.014
Hanáček P., T. Vyhnánek, M. Rohrer, J. Cieslarová and H. Stavĕlíková (2009) DNA polymorphism in genetic resources of red pepper using microsatellite markers. Horticultural Scicience 36:127-132, https://doi.org/10.17221/7/2009-HORTSCI
Hernández V. S. (2014) Importancia del chile silvestre (Capsicum annuum) como recurso genético de México. Mensaje Bioquímico 41:289-303.
Hodgkin T., R. Rana, J. Tuxill, D. Balma, A. Subedi, I. Mar, D. Karamura, R. Valdivia, L. Collado, L. Latournerie-Moreno, M. Saduki, M. Sawadogo, A. H. D. Brown and D. I. Jarvis (2007) Seed systems and crop genetic diversity in agroecosystems. In: Managing Biodiversity in Agricultural Ecosystems. D. I. Jarvis, C. Padoch and H. D. Cooper (eds.). Bioversity International. Columbia University Press. New York, NY. USA. pp:77-116, https://doi.org/10.7312/jarv13648-006
Howard L. R., S. T. Talcott, C. H. Brenes and B. Villalon (2000) Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. Journal of Agricultural and Food Chemistry 48:1713-1720, https://doi.org/10.1021/jf990916t
Ibiza V. P., J. Blanca, J. Cañizares and F. Nuez (2012) Taxonomy and genetic diversity of domesticated Capsicum species in the Andean region. Genetic Resources and Crop Evolution 59:1077-1088, https://doi.org/10.1007/s10722-011-9744-z
IMPI, Instituto Mexicano de la Propiedad Industrial (2010) Declaratoria General de Protección de la Denominación de Origen Chile Habanero de la Península de Yucatán. Diario Oficial de la Federación, Primera Sección. México, D.F. 4 de junio de 2010.
Jones A. G., C. M. Small, K. A. Paczolt and N. L. Ratterman (2010) A practical guide to methods of parentage analysis. Molecular Ecology Resources 10:6-30, https://doi.org/10.1111/j.1755-0998.2009.02778.x
Kalia R. K., M. K. Rai, S. Kalia, R. Singh and A. K. Dhawan (2011) Microsatel- lite Markers: An overview of the recent progress in plants. Euphytica 177:309-334, https://doi.org/10.1007/s10681-010-0286-9
Latournerie M. L., J. L. Chávez S., M. Pérez P., G. Castañón N., S. A. Rodríguez H., L. M. Arias R. y P. Ramírez V. (2002) Valoración in situ de la diversidad morfológica de chiles (Capsicum annuum L. y Capsicum chinense Jacq.) en Yaxcabá, Yucatán. Revista Fitotecnia Mexicana 25:25-33.
Latournerie-Moreno L., J. S. López-Vázquez, G. Castañón-Nájera, J. O. Mijangos-Cortéz, G. Espadas-Villamil, A. Pérez-Gutiérrez y E. Ruiz-Sánchez (2015) Evaluación agronómica de chile habanero (Capsicum Chinense Jacq.). Agroproductividad 8:24-29.
Laurentin H. (2009) Data analysis for molecular characterization of plant genetic resources. Genetic Resources and Crop Evolution 56:277-292, https://doi.org/10.1007/s10722-008-9397-8
Lijun O. and Z. Xuexiao (2012) Inter simple sequence repeat analysis of genetic diversity of five cultivated pepper species. African Journal of Biotechnology 11:752-757, https://doi.org/10.5897/AJB10.2551
Mc Leod M. J., S. I. Guttman and W. H. Eshbaugh (1982) Early evolution of chili peppers (Capsicum). Economic Botany 36:361-368.
Naino Jika A. N., Y. Dussert, C. Raimond, E. Garine, A. Luxereau, N. Takvorian, R.S. Djermacoye, T. Adam and T. Robert (2017) Unexpected pattern of pearl millet genetic diversity among ethno-linguistic groups in the Lake Chad Basin. Heredity 118:491-502, https://doi.org/10.1038/hdy.2016.128
Nei M. (1973) Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences of the United States of America 70:3321-3323.
Nicolaï M., M. Cantet, V. Lefebvre, A. M. Sage-Palloix and A. Palloix (2013) Genotyping a large collection of pepper (Capsicum spp.) with SSR loci brings new evidence for the wild origin of cultivated C. annuum and the structuring of genetic diversity by human selection of cultivar types. Genetic Resources and Crop Evolution 60:2375-2390, https://doi.org/10.1007/s10722-013-0006-0
Oyama K, S. Hernández-Verdugo, C. Sánchez, A. González-Rodríguez, P. Sánchez-Peña, J. A. Garzón-Tiznado and A. Casas (2006) Genetic structure of wild and domesticated populations of Capsicum annuum (Solanaceae) from northwestern Mexico analyzed by RAPDs. Genetic Resources and Crop Evolution 53:553-562, https://doi.org/10.1007/s10722-004-2363-1
Pacheco-Olvera A., S. Hernández-Verdugo, V. Rocha-Ramírez, A. González- Rodríguez and K. Oyama (2012) Genetic diversity and structure of pepper (Capsicum annuum L.) from Northwestern Mexico analyzed by microsatellite markers. Crop Science 52:231-241, https://doi.org/10.2135/cropsci2011.06.0319
Poudel D., B. Sthapit and P. Shrestha (2015) An analysis of social seed network and its contribution to on-farm conservation of crop genetic diversity in Nepal. International Journal of Biodiversity 312621:1-13, https://doi.org/10.1155/2015/312621.
Rohlf F. J. (1998) NTSYS pc: Numerical Taxonomy and Multivariate Analysis System, Versión 2.1. User Guide. Exeter Software. Setauket, New York. 38 p.
Secretaría de Economía (2012) Norma Oficial Mexicana NOM-189- SCFI-2012, Chile Habanero de la Península de Yucatán (Capsicum Chinense Jacq.) Especificaciones y Métodos de Prueba. Diario Oficial de la Federación, Primera Sección. México, D.F. 30 de diciembre de 2012.
SIAP, Servicio de Información Agroalimentaria y Pesquera (2018) Anuario Estadístico de la Producción Agrícola. Servicio de Información Agroalimentaria y Pesquera, SAGARPA. Ciudad de México. http://nube.siap.gob.mx/cierreagricola (Junio 2018).
Sounigo O., R. Umaharan, Y. Christopher, A. Sankar and S. Ramdahin (2005) Assessing the genetic diversity in the International Cocoa Genebank, Trinidad (ICG,T) using Isozyme electrophoresis and RAPD. Genetic Resources and Crop Evolution 52:1111-1120, https://doi.org/10.1007/s10722-004-6110-4
Toledo-Aguilar R., H. López-Sánchez, A. Santacruz-Varela, E. Valadez-Moctezuma, P. A. López, V. H. Aguilar-Rincón, V. A. González-Hernández and H. Vaquera-Huerta (2016) Characterization of genetic diversity of native ‘Ancho’ chili populations of Mexico using microsatellite markers. Chilean Journal of Agricultural Research 7:18-26.
Vera-Sánchez K. S., J. Cadena-Iñiguez, L. Latournerie-Moreno, J. F. Santiaguillo-Hernández, A. Rodríguez-Contreras, F. A. Basurto-Peña, D. Castro-Lara, E. Rodríguez-Guzmán, P. López-López y E. Rios-Santos (2016) Conservación y Utilización Sostenible de las Hortalizas Nativas de México. Servicio Nacional de Inspección y Certificación de Semillas. México, D. F. 132 p.
Yeh F. C., C. Yang and T. Boyle (1999) POPGENE, Version 1.31. Microsoft Window-based Freeware for Population Genetic Analysis. Quick User Guide. University of Alberta and Centre for International Forestry Research. Edmonton, Canada. https://sites.ual-berta.ca/fyeh/popgne.pdf (Enero 2016).
Zizumbo-Villarreal D., P. Colunga-GarcíaMarín, E. Payró C., P. Delgado-Valerio and P. Gepts (2005) Population structure and evolutionary dynamics of wild weedy domesticated complexes of common bean in a Mesoamerican region. Crop Science 45:1073-1083.
Notas de autor