Artículos de revisión
Preservadores maderables y exudados microbianos con actividad antagonista contra agentes biológicos deletéreos
Wood preservatives and microbial exudates with antagonistic activity against biological agents
Preservadores maderables y exudados microbianos con actividad antagonista contra agentes biológicos deletéreos
Revista mexicana de fitopatología, vol. 36, núm. 1, pp. 56-78, 2018
Sociedad Mexicana de Fitopatología A.C.
Recepción: 20 Abril 2017
Aprobación: 30 Septiembre 2017
Resumen: La madera de baja durabilidad es susceptible de deterioro por xilófagos y su protección es una meta tecnológica no alcanzada que genera pérdidas económicas y materiales. Alternativas para evitar el uso de preservadores convencionales tóxicos y contaminantes se pueden establecer mediante el conocimiento del microecosistema bacteriano en la madera. La protección de este material es posible obtenerla con bacterias y sus exudados. El propósito de esta investigación es destacar que el biodeterioro de la madera de baja durabilidad es evitable mediante estrategias microbiológicas relevantes para el control de los hongos xilófagos. La observación es que bacterias de los géneros Arthrobacter, Bacillus y Pseudomonas y sus exudados potencialmente son agentes de protección que exhiben mecanismos de acción diferentes como, el antagonismo, el parasitismo y la producción de exudados que contienen moléculas con efecto: antimicrobiano (antibióticos), quelante (sideróforos) que inhiben enzimas o funciones y compuestos volátiles (dimetilhexadecilamina). El conocimiento de este ecosistema microbiano facilitará la construcción de una alternativa al uso de los preservadores convencionales de madera y constituye un área de oportunidad. Se evidencia la ausencia del conocimiento de bacterias antagónicas de los xilófagos, el manejo de la producción y composición de sus exudados y su aplicación, con lo que se manifiesta un campo inexplorado en la biotecnología de la preservación de la madera.
Palabras clave: xilófagos, lignocelulósico, madera, preservación, durabilidad.
ABSTRACT: Wood of low durability is susceptible of deterioration by xylophages and its protection is a technological goal not reached that generates economic and material losses. Alternatives to avoid the use of toxic preservative conventional and contaminating can be established by knowledge of the bacterial microecosystem in the wood. The protection of this material is possible to obtain it with bacteria and its exudates. The purpose of this research is to highlight that the biodeterioration of low durability wood is avoidable by means of microbiological strategies relevant for the control of xylophages’ fungi. The observation is that bacteria of the genera Arthrobacter, Bacillus and Pseudomonas and their exudates are potentially protective agents that exhibit different mechanisms of action such as antagonism, parasitism and the production of exudates containing molecules with effect: antimicrobial, chelator (siderophores) that inhibit enzymes or functions and volatile compounds (dimethylhexadecylamine). Knowledge of this microbial ecosystem will facilitate the construction of an alternative to the use of conventional wood preservatives and constitutes an area of opportunity. It is evident the absence of knowledge of antagonistic bacteria of the xylophages, the management of the production and composition of their exudates and their application, thus manifesting an unexplored field in the biotechnology of the preservation of wood.
Key words: xylophages’ fungi, lignin, wood, preservation, durability.
Introducción
Por la composición orgánica de la madera es susceptible al deterioro biológico. En general, está constituida por celulosa 40-61%, hemicelulosa 15-30% y lignina 17-35%, particularidad que la hace una excelente fuente de carbono para sostener el ciclo de vida de algunos organismos. La madera en uso está expuesta a organismos tales como, bacterias, algas, hongos e insectos, que al nutrirse de sus macromoléculas provocan su degradación y con ello disminuyen sus atributos físico-mecánicos y prestaciones (Ibáñez, 2012). En la diversidad bacteriana se encuentra un gran número de géneros cuyas especies son capaces de deteriorar la madera al degradar la lignina y modificar su estructura, tales como; Flavobacterium, Xanthomonas, Aeromonas y Nocardia (Singh et al., 2016). También, especies aeróbicas de los géneros de Pseudomonas y Achromobacter igualmente degradan polímeros estructurales, que debilitan la pared celular primaria y secundaria, con la alteración de la estructura cristalina de estos polímeros y los expone para favorecer el desarrollo de otros organismos deterioradores (Zanni, 2004). Las algas xilófagas, están representadas principalmente por especies de los géneros Clorophyta, Chrysophyta y Cianophyta que al deteriorar la madera le provocan un cambio de color, un incremento en la absorción de radiación solar y agua. Sin embargo, el deterioro que realizan las bacterias y algas es menor que el daño producido por hongos e insectos xilófagos, estos últimos representan los principales agentes causantes del biodeterioro (Flower y Gonzalez-Meler, 2015).
En la industria de la construcción y productos basados en madera, los hongos xilófagos son responsables de grandes pérdidas económicas (Arbelo y Garbuyo, 2012). Organismos “cromógenos” se encuentran en este grupo que, al penetrar el tejido leñoso mediante las hifas, alcanzan las cavidades celulares, una acción que tiene como resultado un cambio de color (Moglia et al., 2015). Mientras que las especies fúngicas causantes de “pudrición” se agrupan en diferentes categorías, como las nombradas marrón o cúbica, blanca y suave o blanda. Los hongos se nutren de componentes de pared celular obtenidos mediante enzimas que degradan los polímeros estructurales y con ello alteran las propiedades físico-mecánicas de la madera. La despolimerización de macromoléculas de pared celular la realizan mediante diversas enzimas hidrolíticas que la sintetiza aceleradamente, e.g. la degradación de celulosa cristalina se obtiene con endo-1,4-ß-glucanasas, exo-1,4-ß-glucanasas, 1,4-ß-glucosi-dasas y otras enzimas accesorias (Schmidt, 2007).
Mientras que, para un polímero complejo sin ordenamiento tridimensional como la lignina, conformada por la concatenación de ácidos y alcoholes fenilpropílicos, para su degradación y mineralización el hongo utiliza un complejo enzimático con actividades de oxidasas y peroxidasas. La vía metabólica fúngica para la degradación de este polímero inicia con la acción de la lignina peroxidasa, la cual lisa enlaces carbono-carbono o enlaces éter dentro de la compleja molécula para generar monómeros. Otro complejo enzimático interesante es el que producen algunos basidiomicetos xilófagos que ocasionan la pudrición blanca. Este complejo incluye a la lignina peroxidasa, manganeso peroxidasa, molécula versátil que emplean peróxido de hidrógeno como aceptor de electrones y la lacasa que utiliza al oxígeno molecular. Esta última enzima es una oxidasa constituida por un co-factor con cuatro átomos de cobre, cuyo aceptor de electrones es el oxígeno molecular. También, la lacasa degrada sustratos no fenólicos más difícilmente oxidables en presencia de co-oxidantes; 1-hidroxibenzotriazol (1-HBT), 2,2′-azino-bis-(ácido 3-etilbenzotiazolin-6-sulfónico) o ácido violúrico (Subramanian et al., 2014).
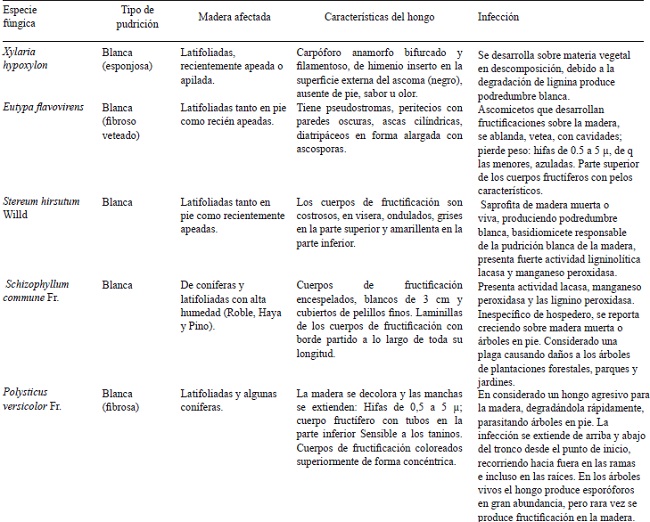
Entre los xilófagos más importantes y considerados perniciosos se tienen a los basidiomicetos, causantes de la pudrición marrón o cúbica (e.g. Serpula lacrymans, Coniophora puteana, Antrodia vaillantii, Lentinus lepideus), capaces de degradar rápidamente la celulosa, hemicelulosa y modificar estructuralmente la lignina. En la pudrición blanca, los basidiomicetos y algunos ascomicetos se nutren preferentemente de lignina, aunque pueden descomponer celulosa. Por la degradación de la lignina y la gran cantidad de celulosa intacta que no utilizan estos hongos, la madera adquiere un color “blanquecino”. Entre las especies fúngicas representativas se encuentra a Trametes versicolor, T. hirsuta y Schizophyllum commune. Mientras que, los ascomicetos causantes de la pudrición suave o blanda (Chaetomium globosum, Monodictys putredinis, Hypocrea muroiana, Cryphonectria parasitica y Fusarium oxysporum), cuyas hifas se desarrollan en el lumen e interior de la pared vegetal secundaria, degradan principalmente celulosa alterando estructuralmente la lignina, con lo cual provocan un ablandamiento característico del material lignocelulósico (Figura 1) (Patel et al., 2013).

Figura 1
Daño provocado por hongos xilófagos en árbol en pie Ganoderma applanatum (a), en la construcción Antrodia vaillantii (b), madera aserrada Ceratocystis sp. (c) y madera derribada Pleurotus ostreatus (d).
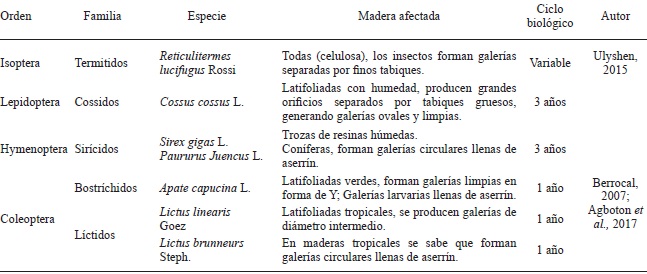
Por otro lado, los insectos constituyen un relevante y diverso grupo dentro del reino animal, con aproximadamente más de un millón de especies descritas y una cantidad de hasta 30 millones de especies aún por conocer (Stork et al., 2015). Asimismo, ellos exhiben una amplia versatilidad para nutrirse y sobrevivir. En relación con el hábito alimentario, los insectos utilizan, e.g. tejido vegetal fresco, materia orgánica en descomposición de origen animal o materia muerta vegetal. Tal es el caso de la madera y los insectos xilófagos, lo hacen mediante la perforación y formación de galerías en la búsqueda de celulosa y almidón. Con ello producen daño físico, mecánico y alteraciones cromáticas en madera almacenada, proceso, servicio y árboles en pie (Cuadro 1) (Zanni, 2008). En este grupo de insectos, destacan los coleópteros (escarabajos o carcomas) y lepidópteros (mariposas y polillas), al igual que isópteros (termitas) e himenópteros (avispas y hormigas). Los isópteros y coleópteros son señalados por producir daños considerables a estructuras y bienes inmuebles de madera (NMX-C-222-1983; Blackwell y d´Errico 2000). También, en gran parte del Norte y Centro América insectos descortezadores del género Dendroctonus (D. frontalis, D. mexicanus) son la principal plaga en bosques de pino, anualmente miles de árboles se pierden por el perjuicio provocado (FAO, 2007).
La protección de la madera de baja durabilidad con preservadores químicos, aunque práctica y económica es ecotóxica, requiere su reemplazo inmediato con preservadores eco-amigables. Por lo que el propósito de este trabajo es establecer las bases de la protección de la madera mediante estrategias microbiológicas.
Clasificación de la madera de acuerdo con su resistencia a xilófagos
La madera puede ser categorizada según el uso y durabilidad (NMX-C-239-1985) y está sujeta a distintos niveles de riesgo y al deterioro por xilófagos. La diferente resistencia presente en este material se debe a diversos factores estructurales y químicos contenidos en el tejido leñoso, variables propias de la especie, edad y condiciones de desarrollo al momento de la tala. De acuerdo con A.S.T.M D-2017-81, la clasificación de la madera por su durabilidad natural se observa en el Cuadro 2A. Esta característica se pondera en años, dicho de otra forma, cuanto tiempo es capaz de mantener sus propiedades mecánicas después del contacto con factores ambientales. Posteriormente, la Junta del Acuerdo de Cartagena (1988) categoriza este material en cinco clases, acorde al deterioro producido por xilófagos (Cuadro 2B). Sin embargo, Bobadilla et al., (2005) catalogan su resistencia conforme al “Criterio de Findlay”, por pérdida de peso después del ataque por hongos (Cuadro 2C) (Findlay, 1951). Este último criterio destaca la importancia del deterioro de la madera causado por hongos xilófagos, así como la relevancia de su control en donde es posible utilizar métodos microbiológicos con tal fin.

Principales preservadores químicos utilizados en madera
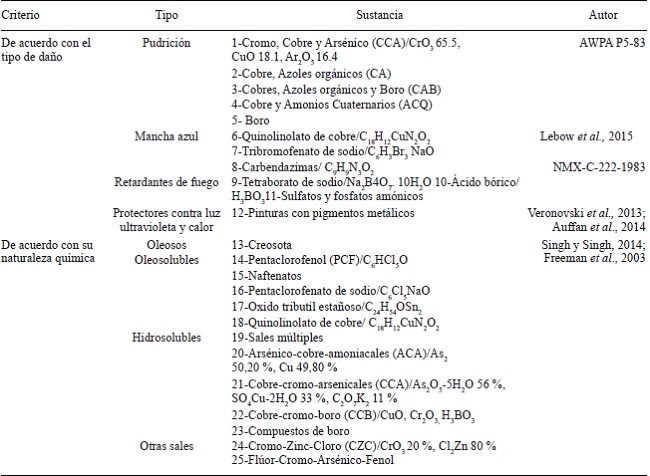
La industria de la preservación química de la madera se desarrolló hasta el comienzo del siglo XIX debido a que ha sido de gran importancia tecnológica, social, histórica y económica, ya que el hombre ha intentado protegerla del deterioro físico y biológico con diversos agentes preservadores. Las mezclas de sales minerales con moléculas orgánicas han sido extremadamente eficaces, donde su clasificación depende de su naturaleza y función (ver el Cuadro 3). Estas sales controlan hongos e insectos en madera en servicio y que está en contacto con el suelo. Su uso se oficializó por la agencia estadounidense protectora de madera (AWPA por sus siglas en inglés) mediante la norma P5-83, tiempo después en México se oficializó (AWPA P5-83). Compuestos como las sales de Cobre-cromo-arsenicales (CCA) son preservadores extremadamente efectivos. Materiales lignocelulósicos tratados con CCA expuestos a condiciones ambientales extremas pueden mantener una vida útil por décadas. Una comprobación realizada por el Departamento de Agricultura de los Estados Unidos de Norteamérica (USDA). Donde se demostró que estacas de madera preservada con CCA, resistieron al ataque de diversos organismos xilófagos por más de 60 años, la USDA predice una vida útil para estos materiales de cinco a diez veces mayor que sin preservar.
La liberación ambiental indiscriminada de estos plaguicidas de uso en la madera ha coadyuvado al alarmante deterioro del ecosistema. Con la finalidad de aumentar la vida media de productos maderables a un mínimo costo económico. Una práctica sociocultural que ahora es un problema ambiental emergente y multifactorial cuya solución es compleja. Sin embargo, en el siglo XX algunas medidas de control para la liberación de plaguicidas se pusieron en marcha en países desarrollados, que tiempo después fueron copiadas y adaptadas en los países subdesarrollados. Una de ellas fue Food Quality Protection Act emitida por el gobierno estadounidense en 1996. En ella, se propuso restringir drásticamente el uso de insecticidas convencionales. Paralelamente, la definición de plaguicida propuesta por la Organización Mundial de la Salud (OMS) se modificó e incluyó a los bioplaguicidas tales como los agentes disuasivos con efecto confusor, inhibidor de la oviposición, anti-alimentario y repelente, así como a los agentes de control biológico (Konradsen et al., 2003). Con estas medidas adoptadas y adecuadas en la legislación mexicana, una nueva oportunidad académica, biotecnológica y comercial se propicia para el diseño de preservadores de origen microbiano para su aplicación en madera de baja durabilidad. El control del biodeterioro de materiales lignocelulósicos mediante preservadores basados en el estilo de vida y estrategias microbianas de invasión ecológica es una alternativa no tóxica.
Compuestos de origen natural con potencial preservador de madera
Hoy en día, métodos de protección con componentes naturales son la alternativa tecnológica para sustituir a los convencionales y nocivos preservadores de la madera de baja durabilidad. La aplicación de esta tecnología eco amigable es lenta y escasamente popular, debido a la practicidad y efectividad a bajo costo económico de los preservadores convencionales.
Dentro de los métodos alternativos en la preservación se exhiben debilidades tecnológicas que se tienen que superar y que son el reto que enfrentar, resolver con rapidez, debido al alarmante deterioro ambiental y al cambio de actitud gubernamental. Entre ellas se incluyen a, las diferencias obtenidas en condiciones controladas (laboratorio) y el rendimiento en campo, las dificultades en eficacia relacionada con la exposición, condiciones ambientales y conflictos en legislación a nivel mundial (Singh y Singh, 2012). Sin embargo, compuestos naturales diferentes, han justificado su eficacia como persevantes de la madera. Históricamente, los extractos de plantas fueron la base molecular para la síntesis química de los piretroides y nicotinoides. Otro ejemplo exitoso de insecticidas son los derivados de los árboles Azadiracta indica (neem) o de Enterolobium cyclocarpum (parota), tales como sus aceites esenciales o componentes contenidos en el duramen como la azadiractina (Martinez et al., 2012; Raya-González et al., 2013). Asimismo, agentes microbianos de control biológico de conocido uso, eficacia y que conforman un paradigma en el control de plagas, tal como Bacillus thuringensis, motiva a conocer el estilo y ciclo de vida de bacterias y otros microrganismos con la finalidad de usarlos en la preservación de la madera. De igual forma, nuevas o novedosas moléculas sintéticas y productos naturales con actividad biológica como controladores de plagas están surgiendo como una solución parcial para el control de organismos deletéreos que dañan la madera (Damian et al., 2010; González-Laredo et al., 2015; Ramírez-López et al., 2016).
La planta sintetiza metabolitos secundarios en la corteza, fruto, hoja, madera, semilla o raíz. Su presencia se evidenciará de acuerdo con su entorno, a sus ciclos circadianos y de vida, donde su función vegetal es variada, entre las que destaca la defensa vegetal y en este arsenal químico vegetal se han descrito compuestos bioactivos inhibidores del crecimiento de patógenos. Al respecto, extractos en hoja de Cinnamomun zeylanicum Ness. contiene componentes que son eficaces para combatir algunos hongos y termitas xilófagas (Cheng et al., 2006; Tascioglu et al., 2013). Recientemente Singh y Singh (2012) reportaron que sustancias de hojas en Elaeocarpus dentatus presentan actividad antifúngica (pudrición parda), de igual forma, aceites esenciales de Citrus x limon, Rosmarinus officinalis, Thymus vulgaris, Cinnamomun zeylanicum Ness. y Syzygium aromaticum, son eficaces para el control de crecimiento en hongos xilófagos productores de moho, los más típicos son especies del género Penicillium, Trichoderma, Fusarium y Aspergillus (Yang y Clausen, 2007; Matan y Matan, 2008) (Cuadro 4). En otros estudios reportados por Kartal et al., (2009), muestran una comparación de formulaciones a base de cinamaldehído, ácido cinámico, aceites de cassia y alquitrán de madera, los cuales inhibieron el crecimiento en Tyromyces palustris y Trametes versicolor, además de proporcionar resistencia contra la termita subterránea Coptotermes formosanus. Autores como Stirling et al., (2007) señalaron que compuestos de Cedrela odorata L. presentan acción biocida, debida a tujaplicina, β-tujaplicinol y ácido plicático, tóxicos en Coniophora puteana, Postia placenta y Trametes versicolor, con características quelantes de metales, simultáneamente el ácido plicático y β-tujaplicinol, mostraron una interesante actividad antioxidante. Tascioglu et al. (2012) reportaron que extractos de corteza (Acacia mollissima) y duramen (Schinopsis lorentzii), mostraron actividad antifúngica debido al gran contenido de taninos condensados, de igual forma, los flavonoides exhibieron una actividad inhibitoria del crecimiento de los hongos Coniophora puteana, Irpex lacteus, Pleurotus ostreatus y T. versicolor (Carrillo-Parra et al., 2011; Nascimento et al., 2013). Se evidencia que las plantas presentan un variado perfil de metabolitos secundarios, que reprimen el crecimiento de una vasta variedad de microorganismos. Sin embargo, el interés se enfoca en la búsqueda de nuevos componentes vegetales para el control de patógenos y se discrimina su importancia en el control de microorganismos deterioradores. Por tanto, la relevancia de los componentes vegetales en la protección de la madera de baja durabilidad se establece como una prioridad de atención inmediata.

Actividad bacteriana antagónica de hongos xilofagos
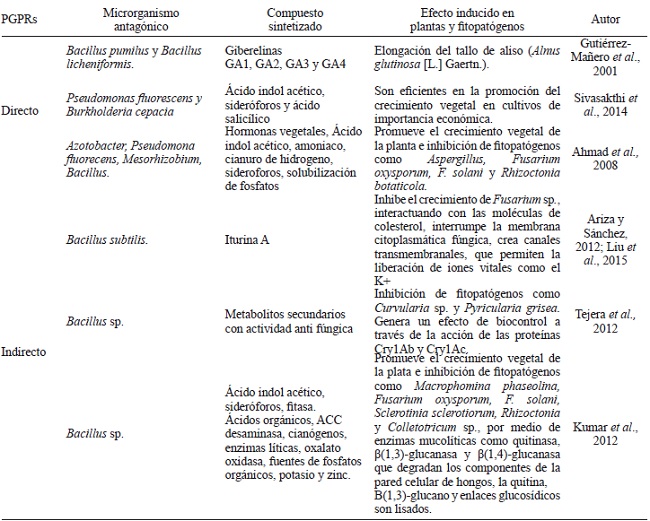
Además de componentes identificados con capacidad preservante de madera, procedentes de plantas como aceites esenciales, ceras, resinas y extractivos, se propone la opción de utilizar a los microorganismos o sus exudados como agentes con actividad antagónica en la protección de la madera contra el deterioro biológico. Del grupo de hongos productores de exudados destacan las especies fúngicas del género Trichoderma, que reprimen el crecimiento de fitopatógenos mediante procesos de micoparasitismo y/o producción de toxinas antifúngicas (Widmer, 2014). También, un amplio grupo de bacterias son capaces de antagonizar el desarrollo de algunos hongos mediante variados mecanismos de acción. En este grupo se encuentran las rizobacterias promotoras del crecimiento vegetal (PGPRs), que por medio de la producción de sideróforos (Saha et al., 2016), antibióticos (Tariq et al., 2017) o competencia por espacios de colonización en raíces y producción de compuestos orgánicos volátiles (COVs), generan un excelente y distintivo efecto antimicrobiano (Cuadro 5).
Los COVs son moléculas orgánicas con masa molecular de <300 Da, se caracterizan por tener una alta presión de vapor que facilita su volatilización (Zou et al., 2007). Algunos tienen potencial antifúngico y pueden actuar a distancia a través de su difusión por el aire e.g. la rizobacteria Arthrobacter agilis UMCV2, en un sistema in vitro limitó el crecimiento del hongo Botrytis cinerea y del oomiceto Phytophthora cinnamomi, debido a la producción de la amina dimetilhexadecilamina (Velázquez-Becerra et al., 2013). Asimismo, con este enfoque experimental Orozco-Mosqueda et al., (2015) caracterizaron la actividad antifúngica de diversas aminas con acción antagónica sobre cuatro cepas de los hongos xilófagos Hypocrea sp. (UMTM3) y Fusarium sp. (UMTM13) obtenidos mediante un escrutinio fúngico en madera en descomposición cuyo origen fue un bosque de pino-encino. El ensayo in vitro evidenció que las aminas evaluadas mostraron un efecto inhibitorio sobre el crecimiento de los aislados fúngicos UMTM, donde destacó la dimetilhexadecilamina. Por su letalidad fúngica es una biomolécula potencialmente útil en el tratamiento preventivo de la madera contra el ataque de hongos xilófagos y de la mancha azul. Dentro de los principales géneros bacterianos identificados como interesantes productores de COVs es Pseudomonas. Ya que Pseudomonas spp. sintetizan ácido cianhídrico (HCN), un inhibidor del crecimiento de patógenos lignocelulósicos. Este metabolito se produce a partir de glicina en condiciones esencialmente microaerofílicas, donde las sintetasas de HCN codificadas por los genes hcnA-BC son fundamentales para la competencia de P. fluorescens (Haas y Défago, 2005). Otras especies bacterianas de este género que producen antifúngicos son: P. chlororaphis (ciclohexanol), P. corrugata (2-etil-1-hexanol), P. aurantiaca (nonanal, benzotiazol y trisulfuro de dimetilo). El compuesto 2,4-Di-ter-burtilfenol lo produce Lysobacter enzymogenes ISE13, utilizado en la represión del ciclo de vida del hongo Colletotrichum acutatum y del oomiceto Phytophthora capsici, ya que inhiben el crecimiento micelial, la esporulación y la germinación de esporas y zoosporas, así como también limitar la formación del apresorio de C. acutatum (Pedraza, 2015). También, un importante número de cepas del género Paenibacillus pueden actuar como antagonistas de hongos fitopatógenos, por ejemplo, P. ehimensis KWN38 sintetiza butanol, el cual genera una inhibición (>50 %) en el desarrollo de hifas de los hongos Rhizoctonia solani, Fusarium oxysporum y del oomiceto Phytophthora capsici (Naing et al., 2014). La bacteria Paenibacillus polymyxa presenta componentes volátiles como 1-Octen-3-ol, benzotiazol, citronelol y 1,3-dicloropropeno, los cuales inhiben severamente el crecimiento micelial y afectan la propagación de patógenos fúngicos, adicionalmente muestran una interesante actividad insecticida y herbicida (Zhao et al., 2011). Asimismo, el efecto de COVs de Streptomyces platensis sobre hongos fitopatógenos se reportó. En dicho estudio se identificaron compuestos antifúngicos como 2-feniletanol y un derivado del felandreno, sustancias responsables de la supresión del crecimiento micelial en Rhizoctonia solani, Sclerotinia sclerotiorum y Botrytis cinerea (Wan et al., 2008). También, cepas de Bacillus subtilis producen COVs que limitan significativamente el crecimiento micelial, producción de pigmentos (inhibición de 43 a 93 % respectivamente) y controlan la germinación de Sclerotinia sclerotiorum, por medio de alquinos, alcoholes, ésteres, cetonas, aminas, fenoles y compuestos heterocíclicos (Liu et al., 2008). Tecnologías microbianas fundamentadas en principios sustentables que emplean agentes de mínima afectación ambiental son la alternativa y a la vez una perspectiva para la preservación de del recurso maderable y derivado.
Estrategias bacterianas para el control de hongos que deterioran la madera
Las bacterias exhiben diversos mecanismos a través de los cuales pueden inhibir el desarrollo de microorganismos causantes del deterioro de la madera y otros productos forestales (Zavattieri et al., 2016; Cely et al., 2016). Alguno de los metabolitos bioactivos bacterianos puede suprimir la germinación, propagación o afectar otras actividades del desarrollo fúngico, tales como la actividad invasora, la supervivencia de los propágulos fúngicos anidados en hendiduras o superficie de la madera para evitar su dispersión (Figura 2). La mayoría de estos bioactivos bacterianos están catalogados en el grupo de antibióticos que inhiben la síntesis de pared celular y la síntesis de proteínas o alterar la estructura del sistema membranal microbiano o provocar la degradación del material genético del microorganismo diana (Maksimov et al., 2011). Algunas especies del género Bacillus producen antibióticos y proteínas insecticidas e.g. B. subtilis producen antibióticos, aunque consideradas PGPRs debido a que ejercen efecto promotor del crecimiento vegetal y un control biológico sobre algunos patógenos del suelo. B. thuringensis produce proteínas que afectan el desarrollo de insectos por lo que se usa como insecticida. Ambos bacilos son una fuente de bioactivos inexplorada para el control de hongos deterioradores de la madera.
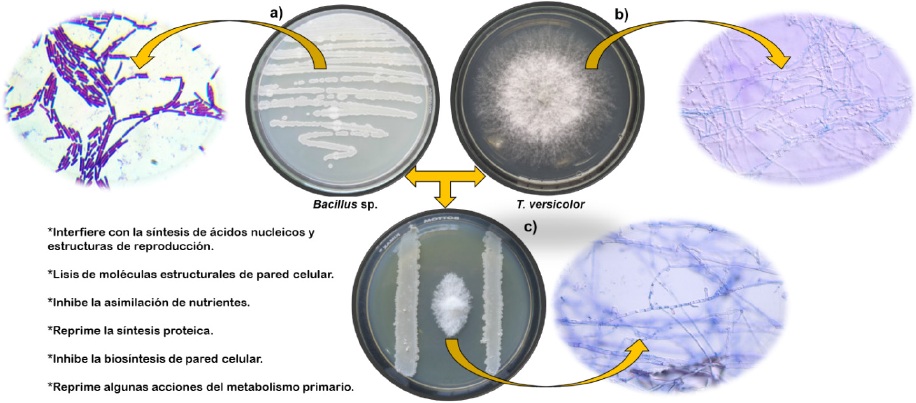
Figura 2
Modelo de inhibición de Trametes versicolor por la secreción de metabolitos de Bacillus sp. (a) el crecimiento del aislado bacteriano de Bacillus sp, (b) ejemplo del desarrollo del hongo fitopatógeno T. versicolor y (c) grado de inhibición de los metabolitos bacterianos sobre el crecimiento del fitopatógeno al estar en desarrollo simultáneo bajo condiciones in vitro a siete días de tiempo.
Los hongos requieren de microelementos para su desarrollo y esta necesidad es de vital importancia. En donde, estos minerales pueden actuar como el blanco o diana farmacológica al tornarlos no disponibles para el hongo y permitir su control. El secuestro molecular de iones evita su disponibilidad y por tanto la asimilación por el hongo y con ello se afecta su desarrollo. Esta estrategia de control fúngico se puede realizar mediante los sideróforos, que son compuestos de bajo peso molecular producidos y secretados por raíces vegetales y algunas bacterias, e.g. los sideróforos capturan elementos como el hierro (Fe+3) (Saha et al., 2016). Las mejores bacterias productoras de sideróforos se encuentran en la categoría bacteriana de las PGPRs. Aunque, los sideróforos son interesantes, por el propósito de este trabajo se muestra como un tema recurrente para su aplicación en la protección de la madera. La capacidad bacteriana de producir sideróforos llama la atención para el secuestro del hierro ya que al formar un complejo Fe3 + sideróforo, e internarlo al citosol bacteriano mediante un receptor específico localizado en la membrana bacteriana, ocasiona que este micronutriente no esté disponible para microorganismos que carezcan del sistema de asimilación específico y reconocimiento de dicho complejo (Radzki et al., 2013; Tariq et al., 2017). De tal manera que al utilizar la totalidad o mayoría del hierro soluble en la madera se suprime el crecimiento fúngico y de otros microorganismos, tal como ocurre en la rizósfera (Bolívar-Anillo et al., 2016; Kumar et al., 2017). La capacidad de los sideróforos para actuar como supresores de hongos deterioradores de la madera se desconoce y es posible que su acción deletérea dependa de factores químicos, físicos y biológicos como ocurre en la rizósfera (Saha et al., 2016). Sin embargo, este es uno de los retos a superar y que puede ser importante en la preservación de este material. Otro campo inexplorado en la preservación de la madera y en el control de su biodeterioro son las enzimas líticas de polímeros de pared celular fúngica. Enzimas que diversos microorganismos las producen entre los que se encuentran las PGPRs, en quienes se han descrito las glucanasas, proteasas y quitinasas. Estas actúan mediante la degradación de los polímeros que componen la pared celular fúngica de los deterioradores de la madera, al igual como se describió que ocurre en hongos fitopatógenos (El-Tarabily y Sivasithamparam, 2006). Entre las principales PGPRs productoras de estas enzimas destacan Bacillus altitudinis y B. amyloliquefaciens (Sunar et al., 2013; Tariq et al., 2017). Como recurso natural renovable pero definitivamente agotable, la madera es de importancia económica a nivel mundial. Un reto en el área de su preservación es tener estrategias basadas en los exudados microbianos para evitar el uso de sustancias tóxicas en su manejo.
Conclusiones
Metabolitos bacterianos como la dimetilhexa-decilamina, 2,4-Di-ter-butilfenol o de naturaleza antibiótica potencialmente son una alternativa a los preservadores convencionales de la madera de baja durabilidad. Pero aún, un desconocimiento existe en cuanto a la producción, modo de empleo y dosificación de los exudados microbianos. Por lo tanto, el reto es alcanzar rendimientos y confiabilidad de estos metabolitos secundarios comparable con los preservadores convencionales mediante estrategias biotecnológicas que incluyen componentes microbiológicos.
Agradecimientos
Los autores agradecen a la UMSNH por el financiamiento a esta investigación.
Literatura citada
Agboton C, Onzo A, Korie S, Tamò M and Vidal S. 2017. Spatial and temporal infestation rates of Apate terebrans (Coleoptera: Bostrichidae) in cashew orchards in Benin, West Africa. African Entomology 25(1):24-36. https://doi.org/10.4001/003.025.0024
Ahmad F, Ahmad I and Khan MS. 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiological Research 163:173-181. https://doi.org/10.1016/j.micres.2006.04.001
Arbelo A y Garbuyo E. 2012. Patologías en la construcción en madera. Estudio de caso: vivienda Punta Colorada. Disponible en línea: https://www.colibri.udelar.edu.uy/bits-tream/123456789/1884/5/ARB6.pdf
Ariza Y y Sánchez L. 2012. Determinación de metabolitos secundarios a partir de Bacillus subtilis efecto biocontrolador sobre Fusarium sp. Nova 10:149-155. Disponible en línea: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S1794-24702012000200002&lng=en&tlng=es
A.S.T.M D-2017-81. American Society for Testing and Materials. (ASTM). 1994. D-2017-81: Standard Method of Accelerated Laboratory Natural Decay Resistance for Woods. Annual Book of ASTM Standard, Philadelphia, v. 410, p. 324-328. Disponible en línea: https://www.astm.org/Standard/standards-and-publications.html
Auffan M, Masion A, Labille J, Diot MA, Liu W, Olivi L, Proux O, Ziarelli F, Chaurand P, Geantetf C, Bottero JY and Rose J. 2014. Long-term aging of a CeO2 based nano-composite used for wood protection. Environmental Pollution 188:1-7. https://doi.org/10.1016/j.envpol.2014.01.016
AWPA P5-83 1983. American Wood-Preserver´s Association (AWPA). 1983. P5-83. Standard for waterborne preservatives. In: American Wood Preservers’ Association AWPA. Pp. 1-4
Berrocal JA. 2007. Clasificación de daños producidos por agentes de biodeterioro en la madera. Revista Forestal Mesoamericana Kurú 4:54-62. Disponible en línea: http://revistas.tec.ac.cr/index.php/kuru/article/view/500/427
Bobadilla EA, Pereyra O, Silva F y Stehr AM. 2005. Durabilidad natural de la madera de dos especies aptas para la industria de la construcción. Floresta 35:419-428. http://dx.doi.org/10.5380/rf.v35i3.5192
Bolívar-Anillo HJ, Contreras-Zentella ML y Teherán-Sierra LG. 2016. Burkholderia tropica una bacteria con gran potencial para su uso en la agricultura. TIP 19:102-108. https://doi.org/10.1016/j.recqb.2016.06.003
Carrillo-Parra A, Hapla F, Mai C y Garza-Ocañas F. 2011. Durabilidad de la madera de Prosopis laevigata y efecto de sus extractos en hongos que degradan la madera. Madera y bosques 17:7-21. Disponible en línea: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S140504712011000100001&lng=es&tlng=en
Cely MV, Siviero MA, Emiliano J, Spago FR, Freitas VF, Barazetti AR, Goya ET, de Souza Lamberti, G, dos Santos IM, De Oliveira AG and Andrade G. 2016. Inoculation of Schizolobium parahyba with Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Increases Wood Yield under Field Conditions. Frontiers in Plant Science 7:1-13. http://dx.doi.org/10.3389/fpls.2016.01708
Cheng SS, Liu JY, Hsui YR and Chang ST. 2006. Chemical polymorphism and antifungal activity of essential oils from leaves of different provenances of indigenous cinnamon (Cinnamomum osmophloeum). Bioresource Technology 97:306-312. http://dx.doi.org/10.1016/j.biortech.2005.02.030
Damian BLM, Martinez MRE, Salgado GR y Martinez PMM. 2010. In vitro antioomycete activity of Artemisia ludoviciana extracts against Phytophthora. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 9:136-142. ISSN:0717-7917. Disponible en linea: http://www.redalyc.org/html/856/85612475009/
El-Tarabily KA and Sivasithamparam K. 2006. Non-strep-tomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biology and Biochemistry 38:1505-1520. http://dx.doi.org/10.1016/j.soilbio.2005.12.017
FAO 2007. Food and Agriculture Organization of the United Nations. State of the world’s forests. Rome, Italy: FAO Forestry Department.
Findlay WPK. 1951. The value of laboratory test on wood preservatives. [S.l.]: Convention British Wood Preserving Association.
Flower CE and Gonzalez-Meler MA. 2015. Responses of temperate forest productivity to insect and pathogen disturbances. Annual Review of Plant Biology 66:547-569. Disponible en línea: http://www.annualreviews.org/doi/pdf/10.1146/annurev-arplant-043014-115540
Freeman MH, Shupe TF, Vlosky RP and Barnes HM. 2003. Past, present, and future of the wood preservation industry: wood is a renewable natural resource that typically is preservative treated to ensure structural integrity in many exterior applications. Forest Products Journal 53(10):8-16. Disponible en línea: http://go.galegroup.com/ps/i.do?id=GALE%7CA110822270&sid=googleScholar&v=2.1&it=r&linkaccess=fulltext&issn=00157473&p=AONE&sw=w&authCount=1&u=umsnh1&selfRedirect=true
González-Laredo RF, Rosales-Castro M, Rocha-Guzmán NE, Gallegos-Infante JA, Moreno-Jiménez MR y Karchesy JJ. 2015. Wood preservation using natural products. Madera y Bosques 21:63-76. http://dx.doi.org/10.21829/myb.2015.210427
Guillén F, Martínez MJ, Gutiérrez A and Del Rio JC. 2005. Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. International Microbiology 8:195-204. Disponible en línea: http://scielo.isciii.es/pdf/im/v8n3/07%20Martinez.pdf
Gutiérrez-Mañero FJ, Ramos-Solano B, Probanza A, Mehouachi J, Tadeo F and Talon M. 2001. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiologia Plantarum 111:206-211. http://dx.doi.org/10.1034/j.1399-3054.2001.1110211.x
Haas D and Défago G. 2005. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nature Reviews Microbiology 3:307-319. http://dx.doi.org/10.1038/nrmi-cro1129
Ibáñez OC, Mantero C, Rabinovich M, Cecchetto G y Cerdeiras P. 2012. Deterioro y preservación de madera. Revista Digital Universitaria 13(5):1-15 Disponible en línea: http://www.revista.unam.mx/vol.13/num5/art55/index.html
Kartal SN, Yoshimura T and Imamura Y. 2009. Modification of wood with Si compounds to limit boron leaching from treated wood and to increase termite and decay resistance. International Biodeterioration and Biodegradation 63:187-190. http://dx.doi.org/10.1016/j.ibiod.2008.08.006
Konradsen F, van der Hoek W, Cole DC, Hutchinson G, Daisley H, Singh S and Eddleston M. 2003. Reducing acute poisoning in developing countries-options for restricting the availability of pesticides. Toxicology 192(2):249-261. https://doi.org/10.1016/S0300-483X(03)00339-1
Kumar H, Dubey RC and Maheshwari DK. 2017. Seed-coating fenugreek with Burkholderia rhizobacteria enhances yield in field trials and can combat Fusarium wilt. Rhizosphere 3:92-99. https://doi.org/10.1016/j.rhisph.2017.01.004
Kumar P, Dubey RC and Maheshwari DK. 2012. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiological Research 167:493-499. http://dx.doi.org/10.1016/j.micres.2012.05.002
Lebow S, Arango R, Woodward B, Lebow P and Ohno K. 2015. Efficacy of alternatives to zinc naphthenate for dip treatment of wood packaging materials. International Biodeterioration and Biodegradation 104:371-376. https://doi.org/10.1016/j.ibiod.2015.07.006
Liu C, Sheng J, Chen L, Zheng Y, Lee DYW, Yang Y, Xu M and Shen L. 2015. Biocontrol activity of Bacillus subtilis isolated from Agaricus bisporus mushroom compost against pathogenic fungi. Journal of Agricultural and Food Chemistry 63(26):6009-6018. https://doi.org/10.1021/acs.jafc.5b02218
Liu W, Mu W, Zhu B and Liu F. 2008. Antifungal activities and components of VOCs produced by Bacillus subtilis G8. Current Research on Bacteriology 1:28-34. Disponible en línea: http://www.docsdrive.com/pdfs/ansinet/crb/2008/28-34.pdf
Maksimov IV, Abizgil’Dina RR and Pusenkova LI. 2011. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens (review). Applied Biochemistry and Microbiology 47:333-345. http://dx.doi.org/10.1134/S0003683811040090
Manual del grupo andino para la preservación de maderas. 1988. Ed. Proyecto subregional de promoción industrial de la madera para construcción de la Junta del Acuerdo de Cartagena L.
Martinez PMM, del Rio RE, Flores GA, Martínez MRE, Ron Echeverria OA and Raya GD. 2012. Enterolobium cyclocarpum (Jacq.) Griseb: The biotechnological profile of a tropical tree. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 11:385-399. Disponible en línea: http://www.redalyc.org/pdf/856/85624131001.pdf
Matan N and Matan N. 2008. Antifungal activities of anise oil, lime oil, and tangerine oil against molds on rubberwood (Hevea brasiliensis). International Biodeterioration and Biodegradation 62:75-78. http://dx.doi.org/10.1016/j. ibiod.2007.07.014
Moglia JG, Amórtegui IC y Giménez AM. 2015. Ocurrencia de la mancha roja en el leño de Aspidosperma quebracho-blanco. Revista de Ciencias Forestales 23:1-2. Disponible en línea: http://www.redalyc.org/pdf/481/48145593003.pdf
Naing KW, Anees M, Kim SJ, Nam Y, Kim YC and Kim KY. 2014. Characterization of antifungal activity of Paenibacillus ehimensis KWN38 against soilborne phytopathogenic fungi belonging to various taxonomic groups. Annals of Microbiolog 64:55-63. http://dx.doi.org/10.1007/s13213-013-06.
Nascimento MS, Santana ALBD, Maranhão CA, Oliveira LS and Bieber L. 2013. Phenolic extractives and natural resistance of wood. Pp:349-370. In Biodegradation-Life of Science. http://dx.doi.org/10.5772/56358
NMX-C-222-1983. Norma Mexicana. “Industria de la Construcción-Vivienda de Madera Prevención de Ataque por Termitas-Especificaciones”.
NMX-C-239-1985. Norma Mexicana para la “Calificación y clasificación de la madera de pino para uso estructural”.
Orozco-Mosqueda M, Valencia-Cantero E, López-Albarrán P, Martínez-Pacheco M y Velázquez-Becerra C. 2015. La bacteria Arthrobacter agilis UMCV2 y diversas aminas inhiben el crecimiento in vitro de hongos destructores de madera. Revista Argentina de Microbiología 47:219-228. http://dx.doi.org/10.1016/j.ram.2015.06.005
Patel N, Oudemans PV, Hillman BI and Kobayashi DY. 2013. Use of the tetrazolium salt MTT to measure cell viability effects of the bacterial antagonist Lysobacter enzymogenes on the filamentous fungus Cryphonectria parasitica. Antonie Van Leeuwenhoek 103(6):1271-1280. https://doi.org/10.1007/s10482-013-9907-3
Pedraza RO. 2015. Siderophores production by Azospirillum: biological importance, assessing methods and biocontrol activity. In Handbook for Azospirillum. Pp. 251-262. Springer International Publishing. 514p. http://dx.doi.org/10.1007/978-3-319-06542-7_14
Radzki W, Mañero FG, Algar E, García JL, García-Villaraco A and Solano BR. 2013. Bacterial siderophores efficiently provide iron to iron-starved tomato plants in hydroponics culture. Antonie Van Leeuwenhoek 104(3):321-330. https://doi.org/10.1007/s10482-013-9954-9
Ramírez-López CB, García-Sánchez E, Martínez-Muñoz RE, Del Río RE y Martínez-Pacheco MM. 2016. Chemical composition of the essential oil from Ageratina jocotepecana and its repellent effect on drywood termite Incisitermes marginipennis. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas. 15(1):53-60. Disponible en línea: http://www.redalyc.org/articulo.oa?id=85643330005
Raya-González D, Martínez-Muñoz RE, Ron-Echeverría OA, Flores-García A, Macías-Rodríguez LI and Martínez-Pacheco MM. 2013. Dissuasive effect of an extract aqueous from Enterolobium cyclocarpum (Jacq) Griseb on the drywood termite Incisitermes marginipennis (Isoptera:Kalotermitidae) (Latreille). EJFA 25:524-530. http://dx.doi.org/10.9755/ejfa.v25i7.15987
Saha M, Sarkar S, Sarkar B, Sharma B K, Bhattacharjee S and Tribedi P. 2016. Microbial siderophores and their potential applications: a review. Environmental Science and Pollution Research 23(5):3984-3999. https://doi.org/10.1007/s11356-015-4294-0
Schmidt O. 2007. Indoor wood-decay basidiomycetes: damage, causal fungi, physiology, identification and characterization, prevention and control. Mycological Progress 6:261-279. http://dx.doi.org/10.1007/s11557-007-0534-0
Schwarze FWMR. 2007. Wood decay under the microscope. Fungal Biology Reviews 21(4):133-170. https://doi.org/10.1016/j.fbr.2007.09.001
Singh AP and Singh T. 2014. Biotechnological applications of wood-rotting fungi: A review. Biomass and Bioenergy 62:198-206. https://doi.org/10.1016/j.biom-bioe.2013.12.013
Singh AP, Kim YS and Singh T. 2016. Bacterial degradation of wood. Secondary Xylem Biology: Origins, Functions, and Applications 9:169-190. https://doi.org/10.1016/B978-0-12-802185-9.00009-7
Singh T and Singh AP. 2012. A review on natural products as wood protectant. Wood Science and Technology 46:851-870. http://dx.doi.org/10.1007/s00226-011-0448-5
Sivasakthi S, Usharani G and Saranraj P. 2014. Biocontrol potentiality of plant growth promoting bacteria (PGPR)-Pseudomonas fluorescens and Bacillus subtilis: A review. African Journal of Agricultural Research 9(16):1265-1277. https://dx.doi.org/10.5897/AJAR2013.7914
Stirling R, Daniels CR, Clark JE and Morris PI. 2007. Methods for determining the role of extractives in the natural durability of western red cedar. International Research Group on Wood Protection. Doc No. IRG-WP 07e20356.
Stork NE, McBroom J, Gely C and Hamilton AJ. 2015. New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods. Proceedings of the National Academy of Sciences 112(24):7519-7523. http://dx.doi.org/10.1073/pnas.1502408112
Subramanian J, Ramesh T and Kalaiselvam M. 2014. Fungal laccases-properties and applications: a review. International Journal of Pharmaceutical and Biological Archive 5(2):8-16. Disponible en línea: http://www.ijpba.info/ijp-ba/index.php/ijpba/article/viewFile/1237/881
Sunar K, Dey P, Chakraborty U and Chakraborty B. 2013. Biocontrol efficacy and plant growth promoting activity of Bacillus altitudinis isolated from Darjeeling hills, India. Journal of Basic Microbiology 55:91-104. http://dx.doi.org/10.1002/jobm.201300227
Tariq M, Noman M, Ahmed T, Hameed A, Manzoor N and Zafar M. 2017. Antagonistic features displayed by Plant Growth Promoting Rhizobacteria (PGPR): A Review. Journal of Plant Science and Phytopathology 1:38-43. Disponible en línea: https://www.heighpubs.org/jpsp/pdf/jpsp-aid1004.pdf
Tascioglu C, Yalcin M, de Troya T y Sivrikaya H. 2012. Termiticidal properties of some wood and bark extracts used as wood preservatives. BioResources 7:2960-2969. Disponible en línea: http://ojs.cnr.ncsu.edu/index.php/BioRes/article/view/BioRes_07_3_2960_Tascioglu_YTS_Termiticidal_Wood_Bark_Extracts
Tascioglu C, Yalcin M, Sen S and Akcay C. 2013. Antifungal properties of some plant extracts used as wood preservatives. International Biodeterioration and Biodegradation 85:23-28. https://doi.org/10.1016/j.ibiod.2013.06.004
Tejera B, Heydrich M y Rojas MM. 2012. Antagonismo de Bacillus spp. frente a hongos fitopatógenos del cultivo del arroz (Oryza sativa L.). Revista de Protección Vegetal 27:117-122. Disponible en línea: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S1010-27522012000200008
Ulyshen MD. 2015. Insect-mediated nitrogen dynamics in decomposing wood. Ecological Entomology 40:97-112. https://doi.org/10.1111/een.12176
Velázquez-Becerra C, Macías-Rodríguez LI, López-Bucio J, Flores-Cortez I, Santoyo G, Hernández-Soberano C and Valencia-Cantero E. 2013. The rhizobacterium Arthrobacter agilis produces dimethylhexadecylamine, a compound that inhibits growth of phytopathogenic fungi in vitro. Protoplasma 25:1251-1262. http://dx.doi.org/10.1007/s00709-013-0506-y
Veronovski N, Verhovšek D and Godnjavec J. 2013. The influence of surface-treated nano-TiO2 (rutile) incorporation in water-based acrylic coatings on wood protection. Wood science and technology 47(2):317-328. http://dx.doi. org/10.1007/s00226-012-0498-3
Wan M, Li G, Zhang J, Jiang D and Huang HC. 2008. Effect of volatile substances of Streptomycesplatensis F-1 on control of plant fungal diseases. Biological Control 46:552-559. http://dx.doi.org/10.1016/j.biocontrol.2008.05.015
Widmer TL. 2014. Screening Trichoderma species for biological control activity against Phytophthora ramorum in soil. Biological Control 79:43-48. https://doi.org/10.1016/j.bio-control.2014.08.003
Yang VW and Clausen CA. 2007. Antifungal effect of essential oils on southern yellow pine. International Biodeterioration and Biodegradation 59:302-306. https://doi.org/10.1016/j.ibiod.2006.09.004
Zanni E. 2004. Patología de madera. Degradación y Rehabilitación de Estructuras de Madera. Primera Edición. Editorial Brujas. Córdoba, Argentina. 244p.
Zanni E. 2008. Patología de la construcción y restauro de obras de arquitectura. Primera Edición. Editorial Brujas. Córdoba, Argentina. 300p.
Zavattieri MA, Ragonezi C and Klimaszewska K. 2016. Adventitious rooting of conifers: influence of biological factors. Trees 30(4):1021-1032. https://doi.org/10.1007/s00468-016-1412-7
Zhao LJ, Yang XN, Li XY, Wei MU and Feng LIU. 2011. Antifungal, insecticidal and herbicidal properties of volatile components from Paenibacillus polymyxa strain BMP-11. Agricultural Sciences in China 10:728-736. http://dx.doi.org/10.1016/S1671-2927(11)60056-4
Zou CS, Mo MH, Gu YQ, Zhou JP and Zhang KQ. 2007. Possible contributions of volatile-producing bacteria to soil fungistasis. Soil Biology and Biochemistry 39:2371-2379. http://dx.doi.org/10.1016/j.soilbio.2007.04.009
Notas de autor
* Autor para correspondencia: cvelazquez@umich.mx.