Artículos científicos
Patrones diurnos del intercambio neto del ecosistema en dos manglares mexicanos de regiones bioclimáticas contrastantes
Diurnal patterns of net ecosystem exchange in two Mexican mangroves from contrasting bioclimatic regions
Patrones diurnos del intercambio neto del ecosistema en dos manglares mexicanos de regiones bioclimáticas contrastantes
Madera y bosques, vol. 30, no. 4, Esp., e3042620, 2024
Instituto de Ecología A.C.
Received: 08 September 2023
Accepted: 11 February 2024
Resumen: Los manglares destacan en los sistemas de carbono azul por su papel en el almacén y captura de carbono atmosférico. Sin embargo, los estudios del intercambio vertical de CO2 son escasos, dejando un vacío en el conocimiento de la dinámica, la variabilidad y los controles ambientales de este flujo. En México, actualmente existen dos sitios monitoreando manglares con la técnica de covarianza de vórtices (eddy covariance), uno en Navopatia (Sonora) y otro en Sisal (Yucatán). En este estudio se utilizaron datos de aproximadamente dos años del intercambio neto de CO2 del ecosistema (del inglés net ecosystem exchange, NEE) y micrometeorología para analizar el comportamiento diurno del NEE durante la época húmeda y la seca. Se empleó un análisis de correlación de Pearson entre el NEE y diversas variables ambientales, así como un Ancova para determinar la significancia de las diferencias observadas. Los resultados mostraron que el NEE en ambos sitios es controlado de diferente manera. En Sonora (clima árido), el ecosistema se comporta principalmente como un sumidero de carbono y sus principales controles son la temperatura, la radiación y el nivel de inundación. Para el manglar de cuenca de Yucatán, los principales controles son el nivel de inundación y el déficit de presión de vapor, comportándose como un neutral sumidero de carbono, en la época húmeda, cambiando hacia una fuente de carbono en la estación seca. Los resultados de este trabajo son importantes para comprender la dinámica de estos ecosistemas costeros de México, así como su respuesta a perturbaciones naturales y antropogénicas.
Palabras clave: Covarianza de vórtices, flujos verticales de CO2, manglar árido, manglar de cuenca, manglar de franja, manglar tropical.
Abstract: Mangroves stand out in blue carbon systems for their role in storing and capturing atmospheric carbon. However, studies of the vertical exchange of CO2 are scarce, leaving a gap in knowing this flux's dynamics, variability, and environmental controls. In Mexico, two sites are currently monitoring mangroves with the eddy covariance technique, one in Navopatia (Sonora) and another in Sisal (Yucatán). In this study, we used data from approximately two years of net ecosystem exchange (NEE) and micrometeorology to analyze the diurnal behavior of NEE during the wet and dry seasons. A Pearson correlation analysis was used between NEE and various environmental variables, as well as an Ancova, to determine the significance of the observed differences. The results showed that NEE at both sites is controlled in different ways. In Sonora (arid climate), the ecosystem behaves mainly as a carbon sink, and its main controls are temperature, radiation, and flooding levels. For the Yucatan basin mangrove, the main controls are flooding level and vapor pressure deficit, behaving as a neutral carbon sink in the wet season, changing to a carbon source in the dry season. The results of this work are important for understanding the dynamics of these coastal ecosystems in Mexico, as well as their response to natural and anthropogenic disturbances.
Keywords: Eddy covariance, vertical CO2 fluxes, arid mangrove, basin mangrove, fringe mangrove, tropical mangrove.
Introducción
A partir del 2009, la Convención Marco de las Naciones Unidas sobre el Cambio Climático (UNFCCC-COP, por sus siglas en inglés) en Copenhague definió colores para el carbono (C), siendo el carbono azul (CA) el relacionado con los manglares, marismas y pastos marinos (Nellemann et al., 2009). Este reconocimiento proviene de la gran capacidad que tienen dichos ecosistemas marinos y costeros para almacenar C especialmente en el sedimento (Alongi, 2020; McLeod et al., 2011). En México se ha impulsado el análisis de la contribución de los manglares a los almacenes de C, especialmente en su distribución tropical (Herrera-Silveira et al., 2016; Herrera-Silveira y Hernández, 2017). Sin embargo, se ha prestado menos atención a flujos ecosistémicos como el intercambio neto de CO2 (NEE) entre el ecosistema y la atmósfera (Alvarado-Barrientos et al., 2021; Granados-Martínez et al., 2021; Vargas-Terminel et al., 2023).
Con la finalidad de evaluar el potencial de mitigación del cambio climático de los ecosistemas como los manglares, es necesario contar con información de almacenes y flujos, así como con las variables biofísicas que los influyen en diferente escala espacial y temporal (Howard et al., 2014; Tarin-Terrazas et al., 2022). El monitoreo directo del intercambio gaseoso entre el ecosistema y la atmósfera se realiza con la técnica de covarianza de vórtices (EC, del inglés eddy covariance) que ha sido ampliamente utilizada en diversos ecosistemas (Baldocchi, 2020). El uso de EC ayuda a caracterizar a escala temporal el NEE, ya que pueden analizarse patrones diurnos, mensuales, estacionales e interanuales. Adicionalmente, se puede asociar el flujo a las condiciones atmosféricas como la temperatura del aire (TA), o el déficit de presión de vapor (VPD), que es una medida de la humedad en la atmósfera y controla el intercambio de gases (Vargas-Terminel et al., 2023).
El NEE del CO2 en los manglares es influenciado por la cantidad de agua dulce que reciben por precipitación (Gnanamoorthy et al., 2020) o sistemas de irrigación (Vargas-Terminel et al., 2023), en general, a escala anual, los aportes de agua dulce incrementan la capacidad de captar CO2 de la atmósfera. Sin embargo, cuando dicho pulso contiene un exceso de nutrientes derivados de actividades como la agricultura se observa un incremento en el proceso de respiración ecosistémica, atenuando la capacidad de mitigar la captura de CO2 (Vargas-Terminel et al., 2023). Por otro lado, se ha observado que durante periodos secos y con altas temperaturas (T > 27 °C), esta capacidad también podría verse disminuida (Alvarado-Barrientos et al., 2021; Liu y Lai, 2019).
Dada la importancia de conocer el NEE de CO2 en los manglares y de realizar estudios comparativos en regiones bioclimáticas contrastantes de México, surgen las preguntas de investigación: ¿cómo cambia la magnitud del patrón diurno del NEE entre un manglar localizado al noroeste del país, específicamente en el estado de Sonora, en comparación con un manglar del sureste mexicano, en el estado de Yucatán, durante la época más húmeda y la más seca? ¿Cómo cambian los distintos controles ambientales en las dos regiones bioclimáticas? Este trabajo está enfocado a responder dichas preguntas.
Objetivos
Comparar el patrón diurno del intercambio neto de CO2 entre el ecosistema y la atmósfera en dos regiones bioclimáticas contrastantes bajo dos distintos periodos estacionales (húmedo y seco) empleando la técnica de covarianza de vórtices.
Materiales y métodos
Sitios de estudio
En el Laboratorio Nacional Conahcyt MexFlux: Monitoreo de Gases de Efecto Invernadero en Ecosistemas, se tienen registrados los observatorios en México en donde hay o hubo mediciones en manglar. Para este trabajo se analizaron los sitios que cumplieran con lo siguiente: (1) Al menos dos años de datos continuos de flujo de CO2, (2) Registro de datos ambientales (temperatura, humedad relativa, radiación, salinidad) e hidrológicos (nivel de inundación), (3) Presentar cambios estacionales en nivel de inundación y cuyas fluctuaciones entre la temporada seca y húmeda sean muy contrastantes (> 20 cm), (4) Regiones climáticas contrastantes con diferente ubicación topográfica (cuenca, franja o petén).
Los sitios que cumplieron estos criterios fueron dos observatorios: 1) El Observatorio de Navopatia, ubicado en Sonora; y 2) el Observatorio Ecohidrológico de Sisal en Yucatán (Fig. 1).
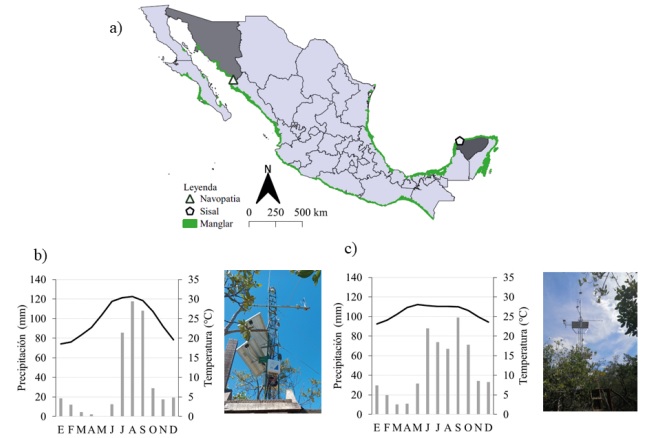
Figura 1
a) Distribución de manglar en México y ubicación de los observatorios de manglar de MexFlux, b) Observatorio ecohidrológico de Navopatia y su climograma (García, 2004) con datos de SMN 26260-DON, SON, 1982-2022, c) Observatorio ecohidrológico del manglar del ejido de Sisal y su climograma con datos de SMN 31029-Sisal, Yuc, 1957-2020.
Observatorio Ecohidrológico de Navopatia
Este observatorio se encuentra al noroeste de México en la bahía de Agiabampo, en la Isla Mazocarit, a 400 m de la estación de Navopatia (Fig. 1 a, b). Este sitio se caracteriza por tener dos especies dominantes de mangle (GranadosMartínez et al., 2021), el negro (Avicennia germinans) y el rojo (Rhizophora mangle). Por otra parte, el régimen intermareal cambia visiblemente a lo largo del año con valores que pueden llegar hasta cerca de los 70 cm sobre la superficie del sedimento (Fig. 2). De acuerdo con datos históricos (1982-2022) de la Estación Don, Sonora (SMN 26260-DON, SON; Servicio Meteorológico Nacional [SMN] y Comisión Nacional del Agua [Conagua], 2024) y de acuerdo con García (2004) el clima es BW(h´)w con una precipitación anual de aproximadamente 400 mm, una temperatura promedio de 30 °C con una mínima de 18 °C y una máxima de 34 °C. La precipitación en la región está influenciada por el Monzón de Norteamérica (NAM) donde la temporada de lluvias comienza en julio y termina en septiembre, mientras que los meses más secos suelen ser abril y mayo (Granados-Martínez et al., 2021).
Observatorio Ecohidrológico del Manglar del ejido Sisal
Este sitio se encuentra al sureste de México, en el Estado de Yucatán, al interior del ejido Sisal (Fig. 1 a, c). El tipo de ecosistema es un manglar de cuenca. Mediciones preliminares en el sitio (Caamal-Sosa et al., 2021) muestran especies de baja altura (dosel promedio 5 m) de Conocarpus erectus (índice de valor de importancia, IVI: 33.3%), Avicennia germinans (IVI: 27.5%), Rhizophora mangle (IVI: 26.4%) y Laguncularia racemosa (IVI: 12.7%). La cantidad de carbono aéreo en árboles y raíces ha sido estimada preliminarmente en 103 Mg ha-1 y 30 Mg ha-1, respectivamente (datos no publicados). La salinidad promedio de la zona es de alrededor de 42 ppt (Caamal-Sosa et al., 2021) y los niveles de inundación máximos rondan los 40 cm sobre el nivel del sedimento (Uuh-Sonda et al., 2023). De acuerdo con los datos (1957-2020) de la estación más cercana, en el poblado de Sisal, Yucatán (SMN 31029-SISAL, YUC; SMN-Conagua, 2024) y a García (2004), el clima es cálido-seco (BSi (h’) W), con un régimen mayoritario de lluvias de verano (precipitación anual promedio de 561 mm) con una contribución de precipitación invernal de entre 5% y 10%. La estación de lluvias puede iniciar entre mayo y junio y terminar en octubre. La temporada de secas transcurre de noviembre a abril, con marzo y abril como los meses más secos. La temperatura media anual es de 26.1 °C, con registros de temperaturas máximas (> 34 °C) en mayo y mínimas (< 18 °C) en enero. La geología de la región es de características calizo-cársticas de gran permeabilidad (Perry et al., 2002).
Monitoreo de flujos y datos biometeorológicos
Para ambos sitios, el sistema de EC consta de un anemómetro sónico 3-D que permite medir la velocidad del viento en sus tres componentes vectoriales (u, v, w), junto con un analizador de gas infrarrojo (IRGA, por sus siglas en inglés) de interfaz abierta, el cual permite medir la concentración de CO2 (así como vapor de agua) en el aire. Conociendo la velocidad del componente vertical del viento, medido con el anemómetro sónico, y la concentración de CO2, obtenido por el IRGA (ambos registrados a una alta resolución temporal: 10 Hz), se puede estimar con precisión el intercambio vertical de CO2 entre la superficie y la atmósfera empleando una técnica estadística conocida como covarianza (Baldocchi, 2003). El anemómetro y el IRGA generalmente se instalan en una torre cuya altura depende del dosel de la vegetación y del área que se intenta investigar. Para el sitio de Navopatia, el sistema de EC se instaló a 1.5 metros sobre la superficie del dosel, mientras que en Sisal este se encuentra a 2.0 metros sobre el dosel.
Junto con los datos de EC, se midieron variables biometeorológicas en el sitio de monitoreo, destacando: temperatura del aire (TA), radiación fotosintéticamente activa (PAR), precipitación (PPT), y nivel de la columna de agua (WL). Las características de los principales instrumentos están enlistadas en la tabla 1. Para el sitio de Navopatia, Sonora, se tienen observaciones de NEE y datos biometeorológicos en el periodo de diciembre de 2017 a diciembre de 2019, mientras que, para el sitio de Sisal, Yucatán, se tienen observaciones de marzo de 2022 a abril de 2023 (Fig. 2), teniendo en ambos sitios, por lo menos un año hidrológico de observaciones. Para la estimación de WL en el sitio de Navopatia hay que considerar que el sensor fue instalado en la parte más baja de la isla que alberga el manglar de franja observado (Granados-Martínez et al., 2021), por lo tanto, la inundación por marea en el sitio de la torre ocurrió únicamente cuando el nivel del agua fue mayor al de esta superficie (nivel de inundación > 0), cubriendo al sedimento. Por el contrario, el nivel del agua por debajo de esta superficie no es registrado, pero se considera exposición de los sedimentos (nivel de inundación = 0). En el caso de Sisal, el sensor de nivel fue instalado al interior de un tubo que funciona como pozo (Uuh-Sonda et al., 2023), con la intención de obtener valores del nivel freático por debajo del sedimento del manglar de cuenca del sitio (nivel de inundación < 0); por lo que un nivel de inundación menor o igual a cero indica una exposición del sedimento.
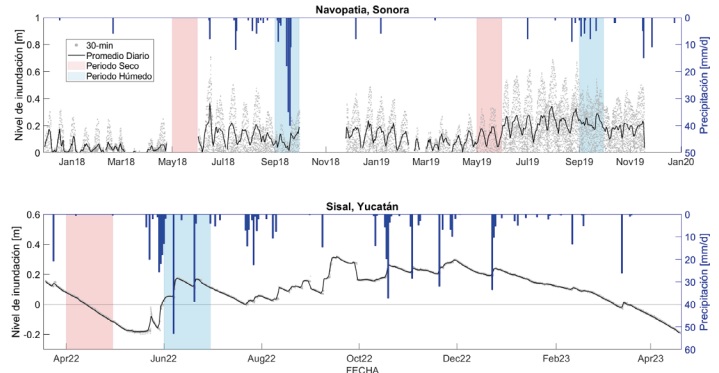
Figura 2
Hidroperiodos para los dos sitios de estudio. Las zonas sombreadas muestran los periodos secos (rojo) y húmedos (azul) identificados para este estudio.
Procesamiento de datos
Para garantizar la calidad y eliminación de datos espurios en los flujos verticales de CO2 (NEE) y los datos biometeorológicos, se realizó en ambos sitios un cuidadoso procesamiento de los datos que constó de dos partes. La primera fue una serie de correcciones estándar a los datos crudos de velocidad de viento y concentración de CO2, siguiendo lo establecido en Foken et al. (2012). Esta serie de procedimientos ayuda a mejorar las estimaciones de NEE a la vez que descarta valores atípicos o poco realistas y corrige problemas propios de la adquisición de datos. Tras estas correcciones, el NEE es estimado para intervalos de media hora, coincidiendo con el valor promedio en el mismo intervalo de los datos biometeorológicos. Esta parte fue realizada con la ayuda del software EddyPro (LICOR Inc., USA). La segunda parte consistió en un control de calidad interno realizado a las estimaciones de NEE y datos biometeorológicos obtenidas en la primera parte. Esta incluyó eliminación de valores atípicos, para lo que se generó una función probabilística de densidad con ajuste gaussiano y mediante un criterio estadístico establecido en dos veces la desviación estándar de todo el conjunto de datos, la eliminación de valores de flujo durante eventos de lluvia y fallas de equipo, y finalmente un filtrado de flujo a velocidades de fricción por debajo de un umbral (0.29 m s-1) donde se presentan condiciones de turbulencia insuficiente. Todas las rutinas de la segunda fase de control de calidad se realizaron en el software MATLAB (The MathWorks Inc., Natick, MA, EE. UU.). Para los datos de Navopatia, Sonora, dicho procesamiento puede ser consultado en Granados-Martínez et al. (2021), mientras que, para los datos de Sisal, Yucatán, el procedimiento es similar a lo mencionado en Uuh-Sonda et al. (2022).
Análisis de datos
En cada sitio se identificó el periodo seco y húmedo de acuerdo con el climograma de cada región y de los datos de precipitación medidos en cada observatorio. Para el Sitio de Navopatia, Sonora, los días con la menor cantidad de precipitaciones se registraron durante el mes de mayo de 2018 y 2019, mientras que aquellos donde se registraron los mayores valores de precipitación ocurrieron en los meses de septiembre de 2018 y 2019. Por su parte, en el sitio de Sisal, Yucatán, se observó que los días con menos precipitación estuvieron en el mes de abril de 2022 y aquellos con la mayor cantidad de lluvias en junio de 2022 (Fig. 2).
Para cada sitio y para cada periodo se calculó el promedio y la desviación estándar a cada media hora del día, para conocer el perfil de comportamiento a lo largo del día y la variabilidad durante todo el periodo, del flujo vertical de CO2 (NEE), y de los principales controles ambientales: radiación fotosintéticamente activa (PAR), temperatura de aire (TA), déficit de presión de vapor (VPD), y nivel de inundación (WL). Para evaluar si las diferencias observadas en los perfiles diurnos del NEE y controles ambientales son estadísticamente significativas, se realizó un análisis de varianza (Andeva; Zar, 2010) a cada par de datos horarios de cada variable en cada sitio. Además, se realizaron ajustes lineales entre el NEE y sus controles ambientales, para comparar el comportamiento entre sitios y periodos. Finalmente, con el fin de evaluar la significancia estadística entre los ajustes lineales realizados, se aplicó un análisis de covarianzas (Ancova; Zar, 2010) a cada par de ellos.
Resultados
Precipitación
Para el sitio de Navopatia, Sonora, durante el periodo seco (mayo 2018 y 2019) no se registraron eventos de precipitación (Fig. 2), como se esperaba para este mes del año (Fig. 1 b). Para el periodo húmedo se registraron 119.0 mm en septiembre 2018 y 37.1 mm en septiembre 2019. Ambos periodos húmedos (septiembre) formaron parte de dos años anormalmente secos en la región, sin embargo, se puede observar que en septiembre de 2018 la precipitación tuvo una magnitud más cercana a la normal (Fig. 1 b), lo cual se debió a la presencia de la tormenta tropical “Sergio” (Granados-Martínez et al., 2021) y no a mecanismos de convección local típicos del NAM, lo cual produjo una inusual entrada de agua dulce al ecosistema. En el caso de Sisal, Yucatán, en el periodo seco (abril 2022) se registró 1.1 mm (Fig. 2), resultando más seco que lo esperado (Fig. 1 c), mientras que en el mes con mayor cantidad de precipitación acumulada (junio 2022) se registraron 117.8 mm, superando la normal para este mes, significando también una importante entrada de agua dulce para el ecosistema.
Perfil diurno del intercambio neto del ecosistema
El NEE es un valor integrado en el tiempo (cada media hora, por ejemplo) del flujo vertical de CO2 que resulta de dos procesos que pueden ocurrir de forma simultánea durante un día en un ecosistema: 1) la asimilación de CO2 por parte de la vegetación, que resulta en un flujo negativo y en un comportamiento de sumidero de CO2 por parte del ecosistema, y 2) la liberación de CO2 o respiración del ecosistema, que resulta en un flujo positivo y en un comportamiento de fuente de CO2. Las figuras 3a y 3b muestran los perfiles diurnos del intercambio neto de CO2 (NEE) entre el ecosistema y la atmósfera para el manglar de Navopatia y Sisal, respectivamente. En ambos sitios, este perfil muestra un patrón esperado, con valores netamente más positivos (alrededor de 10 µmol m-2 s-1) durante las horas de oscuridad o poca luz (de las 17 h 30 a las 6 h 00), puesto que, a falta de luz solar, la respiración del ecosistema es la única componente presente. Por otro lado, en las horas con luz solar (de las 7 h 00 a las 17 h 00), la fotosíntesis (Fig. 3 c, d) comienza a tener relevancia, dominando sobre el proceso de respiración, llevando al flujo de CO2 a valores netamente más negativos (alrededor de -12 µmol m-2 s-1). En ambos sitios, la variabilidad diaria dentro de los dos periodos (seco y húmedo) mostró valores similares de alrededor de ± 3.50 µmol m-2 s-1.
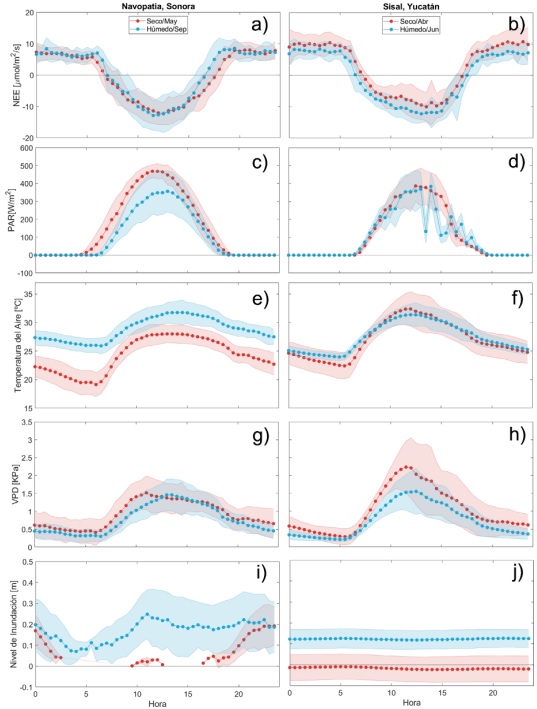
Figura 3
Patrones diurnos en los sitios de Navopatia, Sonora y Sisal, Yucatán, para los periodos seco y húmedo de: a) y b) intercambio neto de CO2 (NEE), c) y d) Radiación fotosintéticamente activa (PAR), e) y f) Temperatura del aire (TA), g) y h) Déficit de presión de vapor (VPD), e i) y j) Nivel de inundación (WL). Las regiones sombreadas indican los momentos en los que ambas distribuciones son estadísticamente diferentes.
Los resultados del Andeva, realizado a los perfiles diurnos del NEE, así como de los controles ambientales, son mostrados en la figura 3, donde las zonas sombreadas muestran los momentos del día en los que los promedios de ambos periodos son estadísticamente distintos. Para el caso del NEE en Navopatia se observa que prácticamente todo el día salvo en ciertos momentos antes del amanecer (de las 5 h 30 a las 6 h 30) y alrededor del atardecer (de las 16 h 00 a las 19 h 00), los perfiles son estadísticamente similares. Por el contrario, en Sisal, los perfiles del NEE entre periodos muestran ser mayormente distintos a lo largo del día.
Controles ambientales
Como se mencionó en la sección anterior, el patrón del perfil diurno del NEE se ve en parte influenciado por el comportamiento de la asimilación de CO2 por fotosíntesis. Por ello, es de esperar que los valores más negativos (un mayor flujo hacia la superficie) del NEE se den cuando los valores de la radiación fotosintéticamente activa (PAR) sean mayores. Las figuras 3c y 3d muestran los perfiles diurnos de la PAR para ambos sitios en ambos periodos. Para el caso del manglar de Navopatia (Fig. 3 c), el periodo seco muestra valores significativamente mayores de PAR y menos variabilidad diaria, comparados con el periodo húmedo. Los máximos de PAR alcanzados alrededor de las 12 h 00 para los días del periodo seco rondan los 467 W m −2 ± 42.83 W m −2 , mientras que, para los días del periodo húmedo alcanzan los 356 W m −2 ± 115.29 W m −2 ,. A diferencia de Navopatia, en el manglar de Sisal, la PAR entre periodos no muestra una significativa diferencia (Fig. 3 d), salvo al amanecer y atardecer, así como durante ciertos momentos después del mediodía (de 13 h 30 a 15 h 30) donde se pueden observar valores más bajos en el periodo húmedo, posiblemente asociados a eventos de nubosidad y precipitación, propios de la temporada. Los valores máximos alcanzados alrededor del mediodía para ambos periodos alcanzan los 385.00 W m-2 ± 83.50 W m-2. Los valores promedio de la PAR para el sitio de Sisal, en ambos periodos, son similares a los observados en el periodo húmedo de Navopatia.
Para ambos sitios, el perfil diario de temperatura del aire muestra el patrón esperado, con las menores temperaturas durante las horas sin luz y los máximos hacia el mediodía. En Navopatia (Fig. 3 e), se observa una marcada diferencia entre las temperaturas registradas en el periodo seco y el periodo húmedo, mostrando en este último, valores más grandes. Para el periodo seco se tienen valores que oscilan entre 19 °C y 28 °C, mientras que en el periodo húmedo oscilan entre 26 °C y 32 °C. La variabilidad diaria para ambos periodos es muy parecida (1.60 °C). En el sitio de Sisal (Fig. 3f) los valores de temperatura del aire entre periodos no muestran una diferencia significativa, salvo en ciertos momentos del día (de 1 h 30 a 7 h 30). Las temperaturas varían entre 22 °C y 32 °C y se observa que existe una mayor variabilidad diaria de la temperatura en el mes más seco.
En ambos sitios, la variación diaria del VPD muestra el patrón esperado con un aumento hacia el mediodía (lo que implica una mayor capacidad de permitir la evapotranspiración dentro del ecosistema) y una reducción en las horas sin luz, similar al comportamiento de la TA. Para el sitio de Navopatia (Fig. 3 g), el perfil diario del periodo seco es ligeramente mayor (atmósfera menos saturada de vapor de agua) al del periodo húmedo, teniendo valores entre 0.30 kPa y 1.50 kPa. Los valores para este sitio varían entre los 0.30 kPa y 1.50 kPa. Por su parte, en Sisal (Fig. 3 h), el VPD del periodo seco es significativamente mayor alrededor del mediodía, que en la época húmeda. Los valores de VPD para el periodo húmedo oscilan entre los 0.20 kPa y 1.56 kPa, mientras que en el periodo seco se observan rangos de 0.30 kPa a 2.23 kPa. Los valores promedios del VPD para el sitio de Navopatia, son similares a los observados en el periodo húmedo de Sisal y similares a los observados en otros manglares (Barr et al., 2010; Leopold et al., 2016; Alvarado-Barrientos et al., 2021).
Los niveles de inundación (Fig. 3i; 3j), en ambos sitios, muestran una marcada estacionalidad, donde los periodos húmedos muestran niveles más elevados que en los periodos secos, que usualmente muestran estar cerca del nivel de sedimento. En cuanto a las magnitudes, los valores máximos en el sitio de Navopatia son aproximadamente 20 cm mayores a los máximos observados en Sisal. La principal diferencia entre ambos sitios radica en la geomorfología de su localización. En el sitio de Navopatia, al ser un manglar de franja, se localiza sobre una isla, siendo la parte más baja de la superficie de esta la referencia del nivel cero (ver Granados-Martínez et al., 2021), así, sus niveles de inundación, y por lo tanto de presencia de agua salada, están modulados principalmente por la variabilidad diaria de la marea (Fig. 2). Durante el periodo húmedo los niveles oscilan entre 0.1 m y 0.25 m, mientras que, en el periodo seco, gran parte del día se tienen niveles muy cercanos a la superficie del sedimento (Fig. 3 i). En el sitio de Sisal, al ser un manglar de cuenca, la presencia de agua en el entorno está más ligada al aumento del nivel freático que varía estacionalmente (Fig. 3) por efecto de las precipitaciones y no de forma diaria, ya que la marea de la zona costera muestra tener muy poca influencia en este sitio (~2 cm), por lo tanto, el nivel cero hace referencia a la superficie de sedimento que cubre las raíces del manglar. Así, durante el periodo seco (abril), el nivel de inundación varía entre 0.05 m y -0.08 m, teniendo el sedimento descubierto la mayor parte del tiempo, mientras que, en el periodo húmedo, los niveles de inundación están por encima del sedimento variando entre 0.07 m y 0.17 m (Fig. 3 j).
Respuesta del intercambio neto del ecosistema a la variación diurna de los controles ambientales
La figura 4 muestra las distintas correlaciones entre la NEE y los controles ambientales aquí estudiados en los dos sitios y en los dos periodos de análisis, a cada media hora del ciclo diurno. Así mismo, la tabla 2 muestra los estadísticos F y p, del Ancova aplicado a cada par de ajustes lineales.
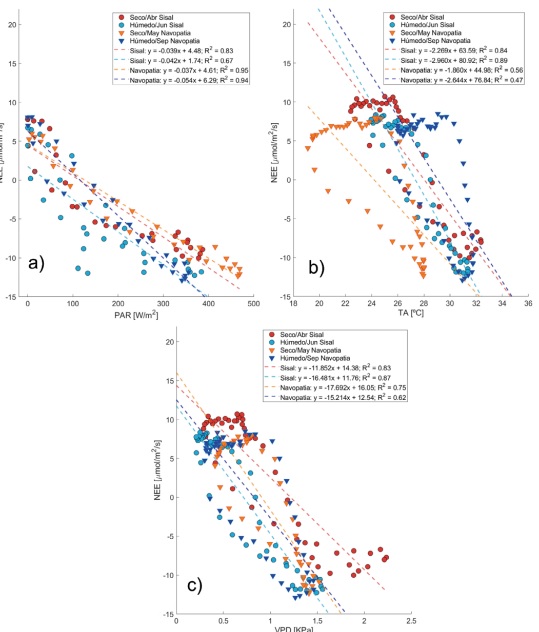
Figura 4
Correlaciones y ajustes lineales, para cada sitio y cada periodo, entre el intercambio neto de CO2 (NEE) y los controles ambientales: a) Radiación fotosintéticamente activa (PAR), b), Temperatura del aire (TA), y c) Déficit de presión de vapor (VPD).
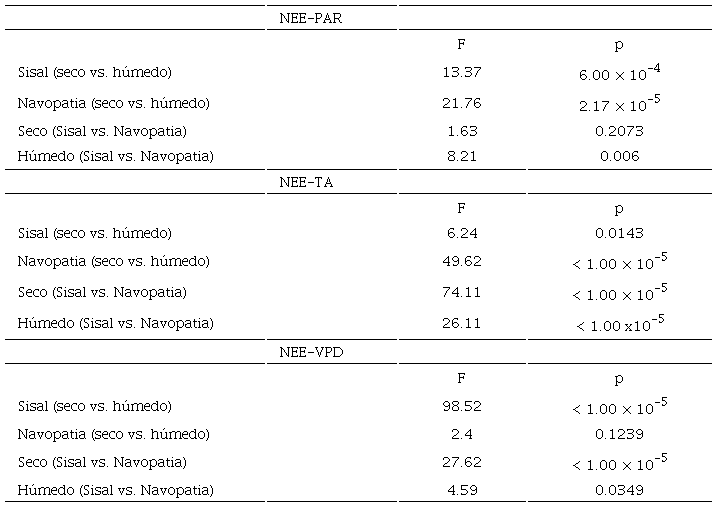
En la figura 4a se puede observar, al comparar en cada sitio el periodo seco contra el periodo húmedo, que las diferencias en humedad ejercen un distinto control en la respuesta del NEE a la variación diurna de la PAR (Sisal: F = 13.37, p = 6.00 × 10-4; Navopatia: F = 21.76, p = 2.17 × 10-5), teniendo la mayor sensibilidad de la NEE a la PAR, para ambos sitios, en los periodos húmedos (comparando las pendientes). Además, al analizar el comportamiento entre ambos sitios, en la época seca, se puede ver que la respuesta de la NEE a la PAR no muestra diferencia estadística (F = 1.63, p = 0.2073), teniendo sensibilidades muy cercanas a los -0.038 µmol s-1 W-1. En cambio, si se observa una diferencia significativa (F = 8.21, p = 0.006), en la respuesta del NEE a la PAR, en el periodo húmedo, donde la mayor sensibilidad es observada en el sitio de Navopatia (-0.054 µmol s-1 W-1 vs. -0.042 µmol s-1 W-1).
Las diferencias de humedad (Sisal: F = 6.24, p = 0.0143; Navopatia: F = 49.62, p < 1.0 × 10-5) también afectan a la respuesta del NEE con la variación diurna de la temperatura del aire (Fig. 4 b). En general, el periodo húmedo de ambos sitios muestra una mayor sensibilidad de la NEE a la TA, comparado con el periodo seco (Sisal: -2.96 µmol m-2 s-1 °C-1 vs -2.27 µmol m-2 s-1 °C-1; Navopatia: -2.64 µmol m-2 s-1 °C-1 vs -1.86 µmol m-2 s-1 °C-1). Y en cuanto a los sitios, el manglar de Sisal muestra ser más sensible a la TA, en cada periodo, que el manglar de Navopatia, pero con un rango de temperaturas no tan grande, como las observadas en el sitio de Navopatia.
Para el control que ejerce la variación diurna del VPD al NEE (Fig. 4 c), se observa que las diferencias de humedad solo afectan al sitio de Sisal (F = 98.52, p < 1.00 x 10-5), ya que para Navopatia no se encontró una diferencia estadística (F = 2.4, p = 0.1239), teniendo una sensibilidad similar. De esta forma, se tiene que la diferencia de sensibilidad es más marcada entre los periodos secos (Sisal: -1 1.85 µmol m-2 s-1 kPa-1; Navopatia: -17.69 µmol m-2 s-1 kPa-1) que, en los húmedos (Sisal: -16.48 µmol µmol m-2 s-1 kPa-1; Navopatia: -15.21 µmol m-2 s-1 kPa-1), de ambos sitios. Siendo la época seca de Navopatia, el periodo con la sensibilidad más alta, seguido del periodo húmedo de Sisal.
Nótese que para el caso de la TA y VPD los puntos (promedios de cada media hora del ciclo diurno) siguen trayectorias que asemejan curvas de histéresis: los máximos negativos de la NEE corresponden al mediodía (cuando la asimilación del ecosistema es máxima), y las condiciones de mañana y tarde son diferentes, provocando que las trayectorias correspondientes difieran.
Discusión
El monitoreo de flujos verticales de carbono mediante la técnica de covarianza de vórtices en manglares ha sido muy limitado y cuenta con muy pocos sitios a escala global (Baldocchi et al., 2018; Gnanamoorthy et al., 2020). Esta tendencia también se refleja en México donde la red de Observacion Mexflux contaba solo con cuatro sitios (Alvarado-Barrientos et al., 2021, Granados-Martínez et al., 2021; Vargas-Terminel et al., 2023) en este tipo de ecosistemas, sin embargo; actualmente sólo dos se encuentran operando.
Los ajustes lineales del NEE a las variaciones diurnas de los controles ambientales (PAR, TA, VPD) muestran cómo este flujo de CO2 es más sensible a las condiciones presentes durante los periodos húmedos que en los secos, es decir, que pequeñas variaciones durante el día de estas variables, en los periodos húmedos, conducen hacia valores más negativos de NEE.
Del análisis a los perfiles diurnos del NEE para ambos sitios (Fig. 3a y 3b) durante el periodo seco, se puede observar que el manglar de Sisal muestra una menor capacidad de asimilación neta de CO2, al poseer magnitudes menos negativas de NEE (de 15.57 a -13.05 µmol m-2 s-1) que el manglar de Navopatia (de 9.61 a -14.76 µmol m-2 s-1). Sin embargo, durante el periodo húmedo ambos ecosistemas muestran una capacidad parecida para asimilar CO2, puesto que presentan valores muy similares de NEE (Sisal: de 12.75 a -15.60 µmol m-2 s-1, Navopatia: de 11.88 a -16.26 µmol m-2 s-1). En este estudio también se observa que el comportamiento diurno del NEE suele mostrar cambios entre la temporada seca y la húmeda, independientemente si el manglar se ubica en una región subtropical, como Sisal, o semiárida, como Navopatia (Tabla 3). Estas diferencias también se han observado en manglares alrededor del mundo. Por ejemplo, en los Everglades, durante las horas de luz en la época seca, se asimilan alrededor de -5 µmol m-2 s-1 menos CO2 que en la época húmeda, mientras que en la noche se emite prácticamente la misma cantidad (Barr et al., 2010). Una comparación de los valores máximos en noche y día del flujo de NEE para cada periodo de este estudio con otros trabajos de la literatura son mostrados en la tabla 3. En el sitio de Sisal se observa que existe una diferencia significativa entre el perfil del periodo seco (cuando los sedimentos están descubiertos) y aquel del periodo húmedo (cuando los sedimentos están inundados). Se puede observar cómo al tener suelos descubiertos (periodo seco) el NEE se vuelve menos negativo a medio día, y más positivo durante la noche. Este comportamiento ya ha sido registrado en otro manglar de cuenca de la Península de Yucatán, en el estado de Quintana Roo (Alvarado-Barrientos et al., 2021). Esto sugiere que la dinámica hidrológica de la Península juega un papel importante en la dinámica de los flujos verticales de CO2 en este tipo de ecosistema. Para el manglar de Quintana Roo también se demostró que las altas temperaturas reducen la capacidad de asimilación del CO2 (Alvarado-Barrientos et al., 2021). Sin embargo, en el sitio de Sisal no se observan diferencias significativas entre las temperaturas de los dos periodos analizados en la mayor parte del día (Fig. 3f). Esto sugiere que la TA no ejercería un control importante sobre la variabilidad estacional del flujo de NEE. El hecho de que, durante el periodo húmedo, este ecosistema sea un mayor asimilador de CO2, probablemente se deba a la disminución del estrés por salinidad por efecto de la entrada de agua dulce (Munns, 2002; Cheeseman y Lovelock 2004; Barr et al., 2009; Alongi y Brickman, 2011; Alvarado-Barrientos et al., 2021). Lo anterior favorece la toma de agua por parte de las plantas y un aumento de su capacidad fotosintética (Leopold, et al., 2016; Lu et al., 2013), considerando que las condiciones de radiación suelen ser similares para los dos periodos del estudio (Fig. 3 d), los valores de VPD durante las horas de luz en este periodo, no se muestran tan extremos (Fig. 3 h) como para reducir la apertura de estomas por tensiones excesivas en el xilema (Grossiord et al., 2020; Gou et al., 2024). Pero también podría deberse a que la respiración de los sedimentos (liberación de CO2 hacia la atmósfera) se ve reducida al propiciar ambientes anóxicos durante los periodos inundados (Alvarado-Barrientos et al., 2021).
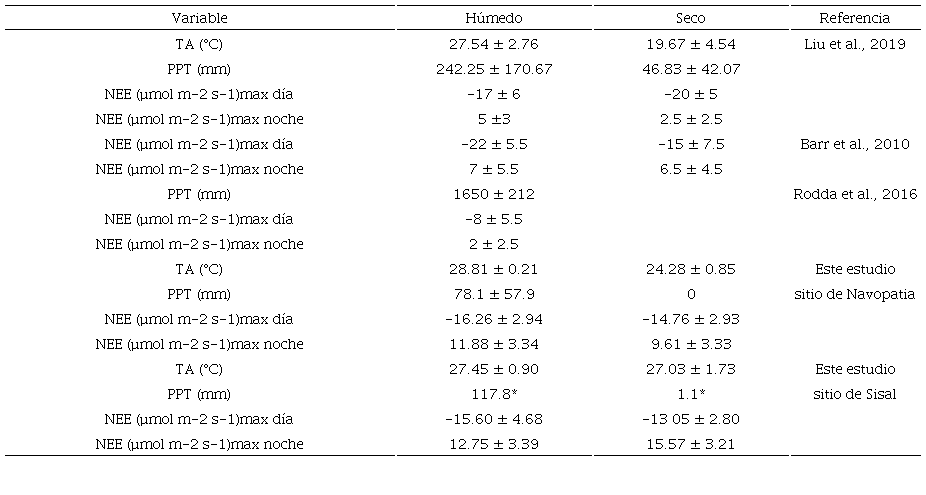
*Acumulado de solo 1 año.
En cuanto al manglar de Navopatia, como se mencionó en los resultados, prácticamente no se observan diferencias significativas del NEE entre ambos periodos, salvo en ciertos momentos del día, siendo los más notorios aquellos que ocurren entre 05 h 30 y 6 h 30 y entre 16 h 00 y 19 h 00. Durante dichos periodos se observan valores de NEE más positivos en el periodo húmedo, lo que podría estar relacionado con las altas temperaturas de aire registradas durante dichas horas (> 31 °C, Fig. 3e). Estudios previos en el sitio de Navopatia, han revelado el control que ejerce la temperatura del aire sobre el comportamiento fenológico del ecosistema, teniendo que en los meses más fríos se observan los máximos de verdor (Granados-Martínez et al., 2021), relacionado a una importante actividad fotosintética. En este contexto, el periodo seco observado (donde se tienen las temperaturas más bajas) presentaría una mayor asimilación de CO2 (NEE más negativos) comparada con el periodo húmedo. También se ha observado que las tasas de respiración del ecosistema aumentan con la temperatura (Alvarado-Barrientos et al., 2021; Xu et al., 2004), comportamiento que también ha sido registrado para este sitio (Granados-Martínez et al., 2021) causando que el perfil de NEE para el periodo húmedo se vuelva más positivo. Sin embargo, a pesar de que existe una marcada diferencia entre las temperaturas del aire, la PAR y el nivel de inundación del periodo húmedo y seco, no se observa una respuesta diferenciable en el comportamiento del NEE (Granados-Martínez et al., 2021) durante el mediodía. Se esperaría que un aumento de la PAR en el periodo seco (Fig. 3 b) causara un aumento en la capacidad fotosintética del ecosistema durante este periodo, haciendo al NEE más negativo a estas horas. Sin embargo, este efecto podría ser contrarrestado con un aumento en la liberación de CO2, debido a que los sedimentos permanecen prácticamente descubiertos durante esta parte del día en este periodo (Fig. 3 i).
Conclusiones
Los resultados sugieren que, bajo condiciones de humedad (periodo con la mayor cantidad de precipitación), tanto el manglar de franja de Navopatia, Sonora, como el manglar de cuenca de Sisal, Yucatán, presentan un comportamiento diurno similar en el intercambio neto de CO2 del ecosistema (NEE), a pesar de que se encuentran en dos regiones bioclimáticamente contrastantes. Sin embargo, para los periodos de secas, el manglar de Sisal muestra un comportamiento de NEE más hacia valores positivos, mientras que en Navopatia la tendencia es mayormente hacia valores negativos. Esta tendencia en Sisal en el periodo húmedo puede ser atribuible a una disminución en el estrés por salinidad debido a las entradas de agua dulce, lo que ayuda al consumo de las plantas, y, por lo tanto, a una mayor actividad fotosintética (a pesar de que la atmósfera se encuentre más saturada de vapor de agua en este periodo, respecto al periodo seco), o por una reducción de la respiración de los sedimentos ya que se propician ambientes anóxicos cuando los sedimentos se encuentran inundados. Por otro lado, la temperatura podría no ejercer control al no existir diferencias significativas de esta entre periodos, siendo la dinámica hidrológica del sitio el principal conductor de estas variaciones. En Navopatia, la tendencia del NEE hacia valores más negativos durante el periodo de secas puede estar relacionada con los decrementos en las temperaturas del aire que favorecen tanto la actividad fotosintética como la disminución de la respiración del ecosistema. Sin embargo, es posible que la exposición de los sedimentos durante la mitad del día en el periodo seco contrarreste la tendencia decreciente del NEE. De esta manera, tanto la dinámica hidrológica del sitio, como las condiciones de temperatura del aire estarían ejerciendo un importante control en este sistema árido.
En la mayoría de los casos, el control que ejercen los cambios diurnos de las variables ambientales (PAR, TA, VPD) sobre el intercambio neto de CO2 del ecosistema (NEE) se muestran diferenciables para cada periodo de estudio (seco y húmedo), siendo el periodo húmedo el que muestra una mayor sensibilidad.
Reconocimientos
Los autores agradecen el apoyo brindado por el M.I. Juan Gómez Liera, técnico académico del Instituto de Ingeniería de la UNAM. Así como las facilidades otorgadas por el ejido Sisal y el M.C. Juan Pablo Caamal Sosa. El primer autor agradece el apoyo ofrecido por la estancia posdoctoral Conahcyt (CVU: 490291) y el apoyo del PROFAPI-ITSON 2023-103. El segundo autor agradece el apoyo del Proyecto CONACYT-SEMARNAT 278608. El cuarto autor agradece el apoyo del PINCC-UNAM 2020 y al proyecto GII No.3117 (subproyecto CA-5) del Instituto de Ingeniería de la UNAM.
Referencias
Alongi, D. M., & Brinkman, R. (2011). Hydrology and biogeochemistry of mangrove forests. En D. F. Levia, D. Carlyle-Moses, & T. Tanaka (Eds.), Forest hydrology and biogeochemistry, synthesis of past research and future directions, ecological studies (pp. 203-219). Springer Netherlands. https://doi.org/10.1007/978-94007-1363-5_10
Alongi, D. M. (2020). Global significance of mangrove blue carbon in climate change mitigation. Sci, 2(3), 67. https://doi.org/10.3390/sci2030067
Alvarado-Barrientos, M. S., López-Adame, H., Lazcano-Hernández, H. E., Arellano-Verdejo, J., & Hernández-Arana, H. A. (2021). Ecosystem-atmosphere exchange of CO2, water, and energy in a basin mangrove of the northeastern coast of the Yucatan Peninsula. Journal of Geophysical Research: Biogeosciences, 126(2), 1-22. https://doi.org/10.1029/2020JG005811
Baldocchi, D. D. (2003). Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future. Global Change Biology, 9(4), 479-492. https://doi.org/10.1046/j.1365-2486.2003.00629.x
Baldocchi, D. D. (2020). How eddy covariance flux measurements have contributed to our understanding of Global Change Biology . Global Change Biology , 26(1), 242-260. https://doi.org/10.1111/gcb.14807
Baldocchi, D., Chu, H., & Reichstein, M. (2018). Inter-annual variability of net and gross ecosystem carbon fluxes: A review. Agricultural and Forest Meteorology, 249, 520-533. https://doi.org/10.1016/j.agrformet.2017.05.015
Barr, J. G., Fuentes, J. D., Engel, V., & Zieman, J. C. (2009). Physiological responses of red mangroves to the climate in the Florida Everglades. Journal of Geophysical Research: Biogeosciences , 114(G2), G02008. https://doi.org/10.1029/2008JG000843
Barr, J. G., Engel, V., Fuentes, J. D., Zieman, J. C., O'Halloran, T. L., Smith, T. J. III, & Anderson, G. H. (2010). Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. Journal of Geophysical Research, 115(G2), G02020. https://doi.org/10.1029/2009JG001186
Caamal-Sosa, J. P., Almonacid-Toledo, A., Esquivel-Campos, R., Sánchez-Mejía, Z., Uuh-Sonda, J. M., Herrera-Silveira, J., Ramírez-Ramírez, G., Valles-Molina, N. C., & Figueroa-Espinoza, B. (2021). Proyecto de conservación voluntaria y mitigación natural del cambio climático en el ejido Sisal, Yucatán. En J. M. Hernández A., M. G. Manzano, M. A. Bolaños, & P. Ibarra (Eds.), Estado actual del conocimiento del ciclo del carbono y sus interacciones en México: Síntesis a 2021. Serie Síntesis Nacionales (pp. 88). Programa Mexicano del Carbono Instituto Tecnológico de Monterrey.
Cheeseman, J. M., & Lovelock, C. E. (2004). Photosynthetic characteristics of dwarf and fringe Rhizophora mangle L. in a Belizean mangrove. Plant, Cell & Environment, 27(6), 769780. https://doi.org/10.1111/j.1365-3040.2004.01181.x
Foken, T., Leuning, R., Oncley, S. R., Mauder M. & Aubinet et al. (2012). Corrections and data quality control. En M. Aubinet, T. Vesala, & D. Papele (Eds.), Eddy covariance: A practical guide to measurement and data analysis. Springer Atmospheric Sci ences. https://link.springer.com/book/10.1007/978-94-007-2351-1
García, E. (2004). Modificaciones al sistema de clasificación climática de Köppen. Universidad Nacional Autónoma de México.
Gnanamoorthy, P., Selvam, V., Deb Burman, P. K., Chakraborty, S., Karipot, A., Nagarajan, R., Ramasubramanian, R., Song, Q., Zhang, Y., & Grace, J. (2020). Seasonal variations of net ecosystem (CO2) exchange in the Indian tropical mangrove forest of Pichavaram. Estuarine, Coastal and Shelf Sci ence, 243, 106828. https://doi.org/10.1016/j.ecss.2020.106828
Gou, R., Chi, J., Liu, J., Luo, Y., Shekhar, A., Mo, L., & Lin, G. (2024). Atmospheric water demand constrains net ecosystem production in subtropical mangrove forests. Journal of Hydrology, 630 130651. https://doi.org/10.1016/j.jhydrol.2024.130651
Granados-Martínez, K. P., Yépez, E. A., Sánchez-Mejía, Z. M., Gutiérrez-Jurado, H. A., & Méndez-Barroso, L. A. (2021). Environmental controls on the temporal evolution of energy and CO2 fluxes on an arid mangrove of Northwestern Mexico. Journal of Geophysical Research : Biogeosciences , 126(7), e2020JG005932. https://doi.org/10.1029/2020JG005932
Grossiord, C., Buckley, T. N., Cernusak, L. A., Novick, K. A., Poulter, B., Siegwolf, R. T., Sperry, J. S. & McDowell, N. G. (2020). Plant responses to rising vapor pressure deficit. New Phytologist, 226(6), 1550-1566. https://doi.org/10.1111/nph.16485
Herrera-Silveira, J. A., Rico, A. C., Pech, E., Pech, M., Ramírez, J. R., & Hernández, C. T. (2016). Carbon dynamics (stocks and fluxes) in Mangroves of Mexico. Terra Latinoamericana, 34(1), 61-72.
Herrera-Silveira, J. A., & Hernández, C. T. (2017). Carbono azul, manglares y políticas públicas. Elementos para Políticas Públicas, 1(1), 43-52.
Howard, J., Hoyt, S., Isensee, K., Telszewski, M., & Pidgeon, E. (Eds.) (2014). Coastal blue carbon: Methods for assessing carbon stocks and emissions factors in mangroves, tidal salt marshes, and seagrasses. Conservation International, Intergovernmental Oceanographic Commission of UNESCO, International Union for Conservation of Nature.
Leopold, A., Marchand, C., Renchon, A., Deborde, J., Quiniou, T., & Allenbach, M. (2016). Net ecosystem CO2 exchange in the “Coeur de Voh” mangrove, New Caledonia: Effects of water stress on mangrove productivity in a semi-arid climate. Agricultural and Forest Meteorology , 223, 217-232. https://doi.org/10.1016/j.agrformet.2016.04.006
Liu, J., & Lai, D. Y. F. (2019). Subtropical mangrove wetland is a stronger carbon dioxide sink in the dry than wet seasons. Agricultural and Forest Meteorology , 278, 107644. https://doi.org/10.1016/j.agrformet.2019.107644
Lu, W., Chen, L., Wang, W., Tam, N. F. Y., & Lin, G. (2013). Effects of sea level rise on mangrove Avicennia population growth, colonization and establishment: evidence from a field survey and greenhouse manipulation experiment. Acta Oecologica, 49, 83-91. https://doi.org/10.1016/j.actao.2013.03.009
McLeod, E., Chmura, G. L., Bouillon, S., Salm, R., Björk, M., Duarte, C. M., Lovelock, C. E., Schlesinger, W. H., & Silliman, B. R. (2011). A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Frontiers in Ecology and the Environment, 9(10), 552-560. https://doi.org/10.1890/110004
Munns, R. (2002). Comparative physiology of salt and water stress. Plant, Cell & Environment , 25(2), 239-250. https://doi.org/10.1046/j.0016-8025.2001.00808.x
Nellemann, C., Corcoran, E., Duarte, C. M., Valdés, L., Young, C. D., Fonseca, L., & Grimsditch, G. (2009). Blue carbon: The role of healthy oceans in binding carbon. United Nations Environment Programme GRID-Arendal. https://portals.iucn.org/library/node/9442
Perry, E., Velazquez-Oliman, G., & Marin, L. (2002) The hydrogeochemistry of the karst aquifer system of the Northern Yucatan Peninsula, Mexico. International Geology Review, 44(3), 191-221. https://doi.org/10.2747/0020-6814.44.3.191
Rodda, S., Thumaty, K., Jha, C., & Dadhwal, V. (2016). Seasonal variations of carbon dioxide, water vapor and energy fluxes in tropical Indian mangroves. Forests, 7(2), 35. https://doi.org/10.3390/f7020035
Servicio Meteorológico Nacional & Comisión Nacional del Agua [SMN-Conagua] (2024). Normales climatológicas por estado. SMN-Conagua. https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado.
Tarin-Terrazas, T., Alvarado-Barrientos, S., Cueva-Rodríguez, A., Hinojo-Hinojo, C., González, E., Sánchez-Mejía, Z., Villarreal-Rodríguez, S., & Yépez, E. A. (2022). MexFlux: sinergias para diseñar, evaluar e informar soluciones climáticas naturales. Elementos para Políticas Públicas , 4(2), 99-118.
Uuh-Sonda, J. M., Figueroa-Espinoza, B., Gutiérrez-Jurado, H., & Méndez-Barroso, L. (2022) Ecosystem productivity and evapotranspiration dynamics of a seasonally dry tropical forest of the northwestern Yucatan Peninsula. JGR Biogeosciences, 127(1), e2019JG005629. https://doi.org/10.1029/2019JG005629
Uuh-Sonda, J. M., Sánchez-Mejía, Z., Figueroa-Espinoza, B. (2023). Análisis de flujos ecohidrológicos ecosistema-atmósfera en un manglar de la Península de Yucatán. En A. Aguilar, E. Yépez, J. García, J. Torres, J. Arreola, R. Barraza, & Z. Sánchez (Comps.). Memorias del VI. Congreso Mexicano de Ecosistemas de Manglar. ITSON. https://www.biodiversidad.gob.mx/media/1/ecosistemas/smmanglares/files/MEMORIAS_VI_CONGRESO_MANGLARES_2023.pdf
Vargas-Terminel, M. L., Rodríguez, J. C., Yépez, E. A., Robles-Zazueta, C. A., Watts, C., Garatuza-Payán, J., Vargas, R., & Sanchez-Mejia, Z. M. (2023). Ecosystem-atmosphere CO2 exchange from semiarid mangroves in the Gulf of California. Journal of Arid Environments, 208, 104872. https://doi.org/10.1016/j.jaridenv.2022.104872
Xu, L., Baldocchi, D. D., & Tang, J. (2004). How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Global Biogeochemical Cycles, 18(4), GB4002. https://doi.org/10.1029/2004GB002281
Zar, J. H. (2010). Biostatistical analysis (5a ed.). Pearson Prentice Hall.
Este documento se debe citar como:
Author notes
* Autor de correspondencia: jorge.uuh.sonda@gmail.com