Dossier
El paso del Pleistoceno al Holoceno en América y el origen de los primeros animales

El paso del Pleistoceno al Holoceno en América y el origen de los primeros animales
Revista Chicomoztoc, vol. 7, núm. 13, pp. 78-101, 2025
Universidad Autónoma de Zacatecas
Resumen: El final del Pleistoceno impulsó importantes adaptaciones en numerosos seres vivos. Una fue la evolución a un ámbito hasta esa época desconocido: el humano. La adaptación de poblaciones animales a territorios alterados por nuestra acción llevó a la formación de la fauna doméstica. Aunque poco se ha estudiado en el continente americano, existen evidencias que avalan la sincronía entre época y origen. En orden cronológico, el perro (Canis lupus familiaris), aunque no se formó en América, fue el primer animal doméstico que ocupó América hace más de 15,000 años. En la zona andina hace unos diez mil años apareció el cuyo doméstico (Cavia porcellus) a partir de Cavia tschudii y la llama (Lama glama glama) del guanaco (Lama glama guanicoe); hace unos ocho milenios, se inicia el proceso con el guajolote (Meleagris gallopavo gallopavo) y hace seis mil años el de la alpaca, derivado de la vicuña (Lama glama vicuña).
Palabras clave: Animales domésticos, continente americano, Pleistoceno, Holoceno.
Abstract: The end of the Pleistocene promoted important adaptations in numerous living beings. One was the evolution to an area unknown until that time: the human. The adaptation of animal populations to territories altered by our action led to the formation of domestic fauna. Although little has been studied on the American continent, there is evidence that supports the synchrony between time and origin. In chronological order, the dog (Canis lupus familiaris), although not formed in America, was the first domestic animal to occupy America more than 15,000 years ago. In the Andean area about ten thousand years ago, the domestic guinea pig (Cavia porcellus) appeared from Cavia tschudii and the llama (Lama glama glama) from the guanaco (Lama glama guanicoe); about eight millennia ago, the process began with the turkey (Meleagris gallopavo gallopavo) and six thousand years ago with the alpaca, derived from the vicuña (Lama glama vicuña).
Keywords: Domestic animals, American continent, Pleistocene, Holocene.
Introducción
Emplear en el mismo enunciado fauna doméstica, América, Pleistoceno, lleva a la interrogante del lector respecto de que se quiere decir, puesto que falsamente se cree que todo tema relevante sobre animales domésticos estaría vinculado con la civilización de Medio Oriente y si hacemos referencia al Pleistoceno, forzosamente pensamos en megafauna, no en cuyos o guajolotes.
¿Existe entonces una razón para abrir un tema así? Ciertamente, y no por ausencia de motivos, sino por la mezcla de falsos conceptos y la percepción de arqueólogos y paleontólogos de que el manejo simultáneo de estos términos lleva a un punto muerto.
Sin embargo, existe suficiente información para asegurar que hay mucho que investigar al respecto, de modo que lo más relevante es modificar nuestra perspectiva y darle un nuevo sentido, algo que no es complicado gracias al enorme avance sobre el tema.
La visión de la domesticación animal en el siglo XXI
En esta época existen dos factores fundamentales para justificar esta nueva visión: en primer lugar tenemos casi tres décadas desde que el tema fue objeto de una visión interdisciplinaria, apareciendo cada vez más estudios de Biología molecular, Bioquímica, Fisiología, Genética, Anatomía, Paleontología, Arqueozoología, Evolución, Zoogeografía, Ecología y otras más, las cuales ofrecen una visión amplia, multidimensional, que lleva a conocimientos sobre este fenómeno que hace medio siglo nadie hubiera imaginado (Valadez, 2021).
Este aspecto es realmente fundamental para comprender la nueva perspectiva que existe. Pensemos un momento. A mediados del siglo pasado todo conocimiento al respecto partía de hallazgos faunísticos lo bastante íntegros para poder ser estudiados, aunque los resultados generalmente llevaban a hipótesis de valor relativo, debido a que la anatomía animal y el contexto eran los únicos medios para entender su presencia, siendo poco probable que pudiera reconocerse algo más. En este momento, basta con muestras de dientes (principalmente) y/o fragmentos de huesos identificados positivamente para hacer múltiples estudios y obtener resultados que reafirman la condición taxonómica, su filogenia y su historia de vida; como ejemplo podemos recordar las múltiples teorías que tuvieron lugar sobre el origen del perro, que iban desde un ancestro ya desaparecido hasta su derivación del chacal, discusión que concluyó cuando los estudios de ADN demostraron que el lobo gris (Canis lupus) era su ancestro inmediato (Vilá et al., 1997: 687; Valadez, 2023: 15-16).
El segundo aspecto refiere a cómo se visualiza en este momento el proceso que llevó a la formación de un animal doméstico. Impulsado quizá por el carácter interdisciplinario mencionado, se considera en el presente que el animal doméstico es producto evolutivo de poblaciones (no especies) que tuvieron la posibilidad de adaptarse progresivamente al ámbito antropógeno gracias a su flexibilidad endocrina y ecológica que permitía obtener beneficios, como tolerar las alteraciones del espacio humano y soportar su propia presencia, a cambio de favorecerse con baja competencia, menos depredación, refugios, alimento y más (Valadez, 2021: 149-165). Dicho proceso no es diferente a los que vemos en casos como las rémoras, los peces piloto y los tiburones, donde se da una forma de interacción en la cual los primeros obtienen beneficios y los segundos mantienen una relación neutra, al menos al inicio y, al paso del tiempo, puede convertirse en algo positivo (Figura 1).
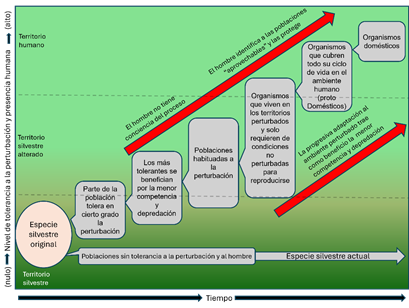
Figura 1
Proceso de domesticación de animales. Durante el proceso, la adaptación progresiva al ámbito humano es fundamental, aunque sólo al final el hombre toma conciencia de esta nueva forma de animal ligada a su espacio (elaborado por Raúl Valadez)
Figura 1. Proceso de domesticación de animales. Durante el proceso, la adaptación progresiva al ámbito humano es fundamental, aunque sólo al final el hombre toma conciencia de esta nueva forma de animal ligada a su espacio (elaborado por Raúl Valadez)
Regresando nuevamente al origen del perro, éste se derivó de la adaptación del lobo chino (Canis lupus chanco). Los estudios del ADN han demostrado de forma indiscutible que desde hace 100,000 años se separaron poblaciones que progresivamente se fueron vinculando con lo humano y modificándose bajo el esquema indicado arriba; de forma que hace 33,500 años tenemos el más antiguo registro fósil de perro en los montes Altai, en el sur de Siberia, mismo que se descubrió en un espacio con mínima presencia de nuestra especie y, hace unos 15,000 años, reconocemos sus entierros específicos en contextos de ocupaciones humanas, mismos que representan las más tempranas evidencias de estos animales como parte de los grupos de cazadores-recolectores (Valadez, 2023: 20-59), lo cual indica que ya en ese momento humanos y perros constituían una sola sociedad (Figura 2). Al margen de lo que es posible reconstruir de esta evolución, es una realidad que se trató de un proceso lento, de forma que se requirieron de más de 66,500 años para pasar por el habituamiento y hasta la protodomesticación; asimismo 15,000 años, o poco más, para ser ya animales ligados al ámbito antropógeno, es decir, ya domésticos.
Más allá de la relevancia de este caso dentro del tema de la domesticación y de su presencia en el continente americano desde hace unos 15,000 años (Cuadro 1), es necesario enfatizar el factor tiempo, pues es un error normal suponer que el paso de lo silvestre a lo doméstico ocurrió en un espacio temporal muy corto, producto del interés humano en apropiarse de ejemplares a los que mantendría cautivos para después promover su reproducción. Así, bajo esta nueva perspectiva, cualquier registro de un animal al que calificamos como doméstico tuvo como precedente un largo periodo de adaptación paulatina a lo humano, casi seguramente desde que hicimos presencia en el continente, hace 20,000 años o poco más.
Planteamiento teórico-metodológico
El objetivo de la presente contribución es presentar los casos de animales domésticos nativos americanos cuyo origen está relacionado con los cambios ambientales y antropógenos ocurridos desde el Pleistoceno superior hasta el Holoceno medio. Para cubrirlo se seguirán los siguientes pasos:
- 1.
1. Presentar datos generales sobre la evolución ambiental en América en el paso Pleistoceno-Holoceno.
2. Mostrar datos generales sobre el desarrollo humano desde el Pleistoceno superior hasta el Holoceno medio en América.
3. Presentar la lista de animales domésticos derivados de la fauna nativa americana según los estudios más recientes, destacando aquéllos cuya evolución en este sentido podemos asociar a los cambios humanos y ambientales ocurridos entre el Pleistoceno terminal y el Holoceno medio.
4. Reconstruir el proceso por el cual se constituyeron poblaciones domésticas de las especies presentadas a partir del paso Pleistoceno terminal-Holoceno y las evidencias de ello.
Pleistoceno-Holoceno en América
Dentro de toda esta dinámica que involucra a la parte animal vale resaltar el aspecto de su flexibilidad ecológica, misma que favorecía a ciertas poblaciones de algunas especies para sobrevivir en diversos ámbitos, no siempre los que esperaríamos. Dicha habilidad, presente en numerosos casos, tuvo una especial relevancia en América desde el Pleistoceno tardío en adelante, debido a los grandes cambios ambientales que tuvieron lugar (Figura 2). En Norteamérica grandes masas de hielo y climas árticos ocupaban, durante la fase glaciar, toda la mitad norte, quedando más al sur la tundra la taiga y estepas; mientras que, en el extremo sur, dominaron los climas templados y subtropicales húmedos o semisecos. Conforme pasó el tiempo y se rebasó la línea de los diez mil años antes del presente (a. p.), al paso del retroceso de los hielos, las temperaturas aumentaron, sustituyéndose gran parte de los bosques templados por praderas y matorrales xerofitos, al tiempo que los bosques tropicales se diseminaron en la parte sur. Estos cambios, ocurridos en tiempos geológicos muy breves de apenas un par de miles de años, llevaron a la extinción a gran cantidad de especies, aunque del mismo modo socorrieron a los que podían ser maleables en sus requerimientos para así sobrevivir en diversos ámbitos, por ejemplo, el antropógeno.
En América del sur las condiciones fueron similares en intensidad, aunque diferentes en la dinámica ecológica. En este caso todas las partes bajas del norte y centro del continente estaban ocupadas por praderas y sabanas tropicales secas, con manchones dispersos de selvas húmedas por aquí o allá. En la parte sur y las zonas altas de los Andes los glaciares dominaban el paisaje y en las zonas intermedias se encontrarían praderas alpinas, pastizales, además de algunas masas de bosques templados (Figura 2). Conforme el Holoceno avanzó, el aumento de temperatura y precipitación llevó a la predominancia de los bosques tropicales perennifolios, sabanas y gigantescos pantanos, quedando diferentes formas de pastizales en los espacios con temperaturas templadas o frías y los hielos perpetuos en las cúspides de las montañas o en el extremo sur del continente. En este proceso la sobrevivencia de las especies dependió también de su flexibilidad para aprovechar toda oportunidad a su alcance.
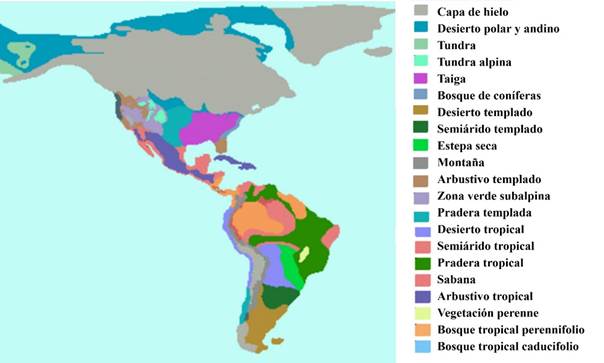
Figura 2
Biomas en el continente americano en la fase glaciar del Pleistoceno tardío (imagen adaptada por Raúl Valadez de https://es.wikipedia.org/wiki/Glaciaci%C3%B3n#/media/Archivo:Ultimo_m%C3%A1ximo_glacial.PNG)
Figura 2. Biomas en el continente americano en la fase glaciar del Pleistoceno tardío (imagen adaptada por Raúl Valadez de https://es.wikipedia.org/wiki/Glaciaci%C3%B3n#/media/Archivo:Ultimo_m%C3%A1ximo_glacial.PNG)
Desarrollo humano en América y la llegada del perro
Según algunos estudios muy recientes, el Homo sapiens pudo haber entrado al continente americano desde hace más de 40,000 años, en plena época glaciar, tal como lo evidencian instrumentos y lascas de roca de 33,000 años de antigüedad recientemente descubiertas en el norte de México (Ardelean et al., 2020: 87-92). Si esta propuesta es correcta, es claro que avanzó sobre una masa terrestre cubierta de hielo, aunque mientras avanzaba hacia el sur encontró territorios de clima más benigno con una fauna abundante y muy diversa. Aparentemente ya hace unos 30,000 años habría llegado hasta Centroamérica.
Durante todo el espacio pleistocénico el ser humano vivió organizado en pequeños grupos denominados “microbandas de cazadores-recolectores (McClung y Zurita, 2014: 265-285)”, desde ese momento quizá el principal cambio que tuvo lugar para con ellos fue la llegada de nuevos grupos que traían consigo a los perros, evento que en México tuvo lugar hace unos 15,000 años (Valadez y Pérez en prensa) (Figura 3).


Figura 3
Restos de perros descubiertos en Rancho Córdova, San Luis Potosí, México. Ésta es la más antigua evidencia de este animal en el continente americano (fotografías de Rafael Reyes, 2015, Instituto de Investigaciones Antropológicas de la UNAM)
Figura 3. Restos de perros descubiertos en Rancho Córdova, San Luis Potosí, México. Ésta es la más antigua evidencia de este animal en el continente americano (fotografías de Rafael Reyes, 2015, Instituto de Investigaciones Antropológicas de la UNAM)
En esta región del mundo, el final del Pleistoceno marca también el retroceso del hielo e, igualmente, la desaparición de gran parte de la fauna de mamíferos y aves, en especial de la llamada “megafauna”; coincidencia que caracteriza la evolución ecológica de Norteamérica, mientras que en otros continentes las extinciones antecedieron a los cambios ambientales. De esta forma podemos considerar el periodo ubicado entre los diez y nueve mil años antes del presente como un momento clave, tanto por el progresivo aumento en la temperatura como por la notable pérdida de la diversidad faunística, acaso producto no sólo de los efectos climáticos sino también del impacto de esta nueva versión de las microbandas constituidos por Homo sapiens y Canis lupus familiaris (Valadez, 2023: 57-59).
Este conjunto de modificaciones necesariamente provocó cambios en las formas de subsistencia de los grupos humanos. ¿Qué relación podemos encontrar entre esto y los procesos que llevaron a la domesticación? En las porciones más septentrionales, en donde los hielos y la tundra fueron sustituidos por estepas y taigas, el esquema biológico fue similar al del pasado, pues además de los climas fríos y muchas veces extremos, ahí encontramos gigantescos rebaños de bisontes (Bison bison), grandes poblaciones de borregos y cabras (Ovis canadienses y Oreamnos americanus), de berrendos (Antilocapra americana) y de diversos cérvidos (Odocoileus virginianus, Odocoileus hemionus, Cervus elaphus, Cervus canadiensis, Rangifer tarandus y Alces alces). Todo ello llevó a que la forma de vida humana no sufriera modificaciones profundas y, así, los pequeños grupos nómadas de cazadores-recolectores subsistieron por muchos milenios más (Valadez, 2023: 178-180).
Respecto de los grupos que habitaban las regiones más al sur, éstos vivieron bajo condiciones ecológicas muy diferentes que promovieron nuevas formas de aprovechar los recursos. En un marco muy general la porción sur de Norteamérica, es decir, México y Centroamérica, se encuentra mayormente inmersa en la llamada zona tórrida o tropical y con costas, a veces muy próximas, entre los océanos Pacífico y Atlántico, lo cual, en primera instancia, involucra un clima húmedo y cálido. Se considera que los grupos humanos que fueron llegando al territorio mexicano siguieron los cursos de costas y ríos para explorar nuevos territorios. Ya en el centro y sur de México se encontraron con ríos de caudal más abundante y continuo, así como con ricas masas lacustres que les permitían asentarse por periodos más largos.
Gracias a ello, a partir del Holoceno temprano, el modo de vida de las bandas humanas va cambiando en la mayor parte del continente hacia un sistema de organización llamado “macrobandas estacionales” (McClung y Zurita, 2014: 265-270) que permanecían tiempos mayores en zonas donde los recursos y el clima eran más favorables. Esta nueva forma de organización derivó en que ya desde los ocho mil años en adelante aparecen evidencias de plantas que son aprovechadas y seleccionadas por estos grupos y un par de miles de años después el cultivo de plantas es una realidad.
Para el caso sudamericano las condiciones fueron diferentes. Durante el final del Pleistoceno tuvo lugar también la extinción de la megafauna, quizá causada, o al menos favorecida, por ambientes extremos que, por un lado, promovieron el desarrollo de una cuenca amazónica en la que los cauces de ríos y gigantescos humedales y pantanos limitaron la existencia de poblaciones como perezosos gigantes, proboscídeos, gliptodontes y otros más, los cuales no podían adaptarse a estos ambientes; pero, por otro lado, al mismo tiempo las regiones meridionales y la andina avanzarían hacia climas más áridos, condición que tampoco asistiría su existencia. Respecto de las poblaciones humanas que poco a poco fueron adaptándose a estos cambios, vale tomar en cuenta de forma especial a las que ocuparon los Andes, pues fue una región clave en el desarrollo de las formas domésticas (Valadez, 2023: 270-271). Esta región, en el final del Pleistoceno, estaba cubierta por praderas, mismas que al paso del Holoceno se hicieron menos frías pero más y más secas, convirtiéndose en lo que en el presente se denomina “puna”. Ahí los grupos humanos continuaron con su vida nómada por muchos siglos más, cazando guanacos (Lama glama guanicoe) y las diferentes especies de venados presentes, además de aprovechar los escasos recursos vegetales. Sólo en tiempos de lluvia se establecían en mayor cantidad en las cañadas y lechos de arroyos, explotando todo lo posible (Valadez, 2023: 270-271; en prensa).
Fauna doméstica nativa de América y el inicio del Holoceno
Con base en lo anterior, conforme va teniendo lugar el final del Pleistoceno en el continente americano, nos encontramos con ambientes cambiantes que ponen a prueba a todas las formas de vida. Respecto de la fauna, en el presente es relativamente fácil encontrar evidencia de especies que hace 10,000 años tuvieron una distribución muy diferente a la actual; por ejemplo, camélidos que existían en México y poco a poco fueron limitando su existencia a la zona andina, o bien casos en los que varias de sus poblaciones las encontramos ahora en reductos, refugios ecológicos, donde han logrado sobrevivir al paso de los siglos a muchos miles de kilómetros de donde se encuentran las demás: uno de estos casos es el de diversos cricétidos propios de climas templados y fríos, pero que actualmente existen en zonas montañosas del centro de México o las sierras del estado de Chiapas (Hall, 1981: 740-747).
Este laberinto ecológico fue también el promotor de que en algunos casos las poblaciones de ciertas especies aprovecharan la presencia humana, o quizás debieron entrar a un periodo de tolerancia, ya que no existían alternativas, colaborando con ello a la entrada al proceso que derivaría primero en el habituamiento al entorno humano, para posteriormente introducirse más y más en él conforme el balance entre beneficios e inconvenientes se inclinaba a favor de la interacción.
En la Figura 4 tenemos una síntesis de los principales animales que dieron lugar a formas domésticas en este continente (Valadez, 2023: 177-306). Como puede verse, los momentos que dieron inicio al proceso son muy variables, pero para nuestros objetivos los casos a considerar son los que se vinculan con el final del Pleistoceno, el Holoceno temprano y los cambios en el modo de vida de los grupos humanos durante esta segunda fase, con el subsecuente impacto en los animales involucrados.
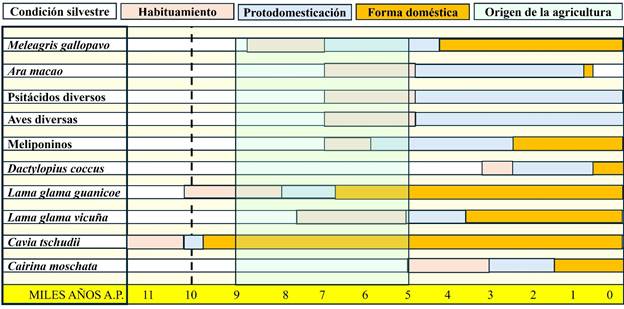
Figura 4
Fauna americana relacionada con la domesticación. La línea punteada representa el final del Pleistoceno, momento a partir del cual tuvieron lugar cambios ambientales que promovieron modificaciones en los esquemas de vida humanos y de diversos animales que aprovechaban los beneficios que obtenían (figura elaborada por Raúl Valadez)
Figura 4. Fauna americana relacionada con la domesticación. La línea punteada representa el final del Pleistoceno, momento a partir del cual tuvieron lugar cambios ambientales que promovieron modificaciones en los esquemas de vida humanos y de diversos animales que aprovechaban los beneficios que obtenían (figura elaborada por Raúl Valadez)
Reconstrucción de los procesos hacia la domesticación en el Pleistoceno y Holoceno temprano
El cuyo (Cavia tschudii)
El más temprano caso de un animal americano que se involucró con la domesticación fue el cuyo, al que se le reconoce como Cavia porcellus, aunque el ancestro silvestre ha sido objeto de discusión. Las evidencias arqueozoológicas ubican su condición doméstica desde finales del Pleistoceno, es decir, antes de cualquier cambio en las formas de vida humana, incluso antes de que las grandes extinciones tuvieran lugar.
Uno de los espacios arqueozoológicos con evidencia al respecto se ubica en la sabana de Colombia (Lord et al., 2020), específicamente en abrigos rocosos del sitio de Tequendama sobre los nueve mil años a. p., en el cual se reconocieron miles de restos de cuyos. La propia cantidad y su morfología ósea llevó a la conclusión de que se trataba de individuos domésticos y que, por tanto, éste habría sido su centro de origen (Altamirano, 1986: 22-29) (Figura 5). Sin embargo, estudios recientes del ADN de 12 muestras arqueozoológicas mostraron que no pertenecían a la forma doméstica actual (Lord et al., 2020) y que la especie silvestre con más afinidad en este sentido era C. anolaimae.
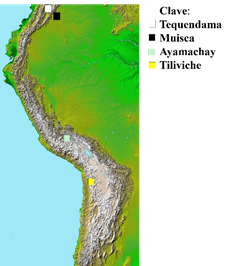
Figura 5
Mapa con los restos más antiguos de cuyos y cuadro con temporalidades y contextos (Valadez, 2023: 297-302; mapa de Nasa Anden - civilizaciones andinas - wikipedia, la enciclopedia libre, adaptado por Raúl Valadez)
| Sitios | Antigüedad probable (años a. p.) | Contexto | Evidencia de domesticación | Posibles usos |
| Tequendama, Colombia | 9000 | Basureros | Cantidad | Alimento |
| Muisca, Altiplano Cundiboyascence, Colombia | 9400-8440 | Basureros | Cantidad | Alimento |
| Ayamachay, puente Ayacucho, Kotosh, Perú | 9000-2500 | Basureros | Intensidad de uso, osteometría | Alimento |
| Tiliviche, Chile | 10000-6000 | Doméstico | Ejemplares fuera de área de distribución | Alimento, ¿ritual? |
Figura 5. Mapa con los restos más antiguos de cuyos y cuadro con temporalidades y contextos (Valadez, 2023: 297-302; mapa de Nasa Anden - civilizaciones andinas - wikipedia, la enciclopedia libre, adaptado por Raúl Valadez)
Otras investigaciones en la misma región, en abrigos rocosos de El Abra y el Altiplano Cundiboyascence (Figura 5), han llevado también a la conclusión de que en esa zona desde épocas muy tempranas tuvo lugar un proceso de intensa interacción ser humano-cuyos, derivando en poblaciones que, al paso del tiempo, quedaron constituidas por ejemplares más grandes, de mayor peso, con modificaciones en el dentario y pelvis, lo cual se interpretó como evidencia de cambios involucrados con su domesticación (Ijzereef, 1978: 165-168).
En la sierra central del Perú (Figura 5), en los sitios de Ayamachay, Puente de Ayacucho y Kotosh (Rofes y Wheeler, 2003: 29-45) se han reconocido sitios de antigüedad semejante a los colombianos con evidencias relacionadas con Cavia porcellus. En éstos la afirmación se basó en pruebas que se van encontrando en función de la antigüedad, de modo que en los contextos más añejos (9,000 años a. p.) los restos aparecen en gran cantidad dentro de la basura doméstica, quemados y con huellas de corte, manifestando así su empleo como alimento; reconociéndose en los restos ubicados sobre los 5,500 años a. p. (Ayamachay y Puente de Ayacucho) una alta variabilidad osteométrica, sobre todo de los dentarios, lo cual se interpretó como evidencia de que se trataba de poblaciones cuya variabilidad dependió de los intereses humanos.
También tenemos el caso del sitio de Tiliviche en el norte de Chile, un lugar ubicado en la montaña a una altura media (950 msnm), con una antigüedad entre los diez mil y seis mil años a. p. (Hesse, 1984) (Figura 5). Aunque la alimentación de los pobladores dependía fundamentalmente de la pesca y la colecta de moluscos marinos (81 por ciento del total de fauna estudiada), se reconoció una muestra de 62 especímenes (0.9 por ciento) de Cavia, presumiblemente C. porcellus, ya que no existe este género en forma silvestre. Aunque la cantidad de elementos no son muchos, fue el único roedor reconocido para este momento, observándose además algunas peculiaridades, por ejemplo que los huesos se encontraron dispersos de una manera uniforme en el área de excavación y el predominio de cráneos y dentarios en la muestra (42 de 62 especímenes), todo lo cual fue interpretado como el resultado de una intencionalidad, tanto en lo que refiere a la selección de partes a usar como de la forma en que dichas piezas fueron colocadas.
A partir de diversas muestras arqueozoológicas se realizaron estudios del ADN y los resultados mostraron que su ancestro silvestre fue C. tschudii. De esta forma en el presente se considera que C. porcellus es su derivado doméstico actual, aunque cabe saber si este roedor pasó a la condición doméstica en más de una ocasión.
El género Cavia se distribuye en buena parte de Sudamérica, con excepción de la cuenca del Amazonas y el extremo meridional del continente. Se le encuentra desde zonas costeras hasta los 4,500 metros sobre el nivel del mar. Ocupa ambientes secos y alpinos, pero cercanos a fuentes de agua; preferentemente entre matorrales densos. Su alimentación es completamente herbívora, de vida nocturna, conforma pequeños grupos, ocupa cavidades excavadas. Cuando se siente en peligro se queda inmóvil y a la primera oportunidad se escabullirá rápidamente. Ocupa zonas de cultivo sin problemas (Anónimo, 2018).
Sin duda la aparición de poblaciones domésticas, aparentemente derivadas de más de una especie, fue producto de su habilidad para aprovechar los espacios humanos en condición de comensal, mucho antes de que surgiera el cultivo de plantas. Parece factible que conforme el Homo sapiens se dispersó por Sudamérica, este roedor, de forma equivalente a como lo hicieron Mus y Rattus en Asia, se beneficiarían del ámbito humano; con la diferencia de que no se convirtieron en competidores del hombre ni transmitían enfermedades. De esta forma, desde el Pleistoceno superior los cuyos pasarían a la fase de habituamiento en diversos lugares, utilizando los campamentos humanos como refugio, como madrigueras, como lugares para hacer sus nidos y alimentándose de la vegetación aledaña y de lo que el hombre desechaba. Su presencia tenía un mínimo impacto en los espacios humanos con la ventaja de que se le podía atrapar con facilidad. Así, y bajo un esquema de que unos y otros se favorecían por la relación, para inicios del Holoceno ya existía como organismo doméstico. Las razones por las cuales sólo C. tschudii continuó el proceso bajo el nombre de C. porcellus es algo que aún no podemos determinar, quizá mayor habilidad para aprovechar el ámbito antropógeno, mayor capacidad reproductiva o más fácil manejo por parte de las personas (Figura 6).
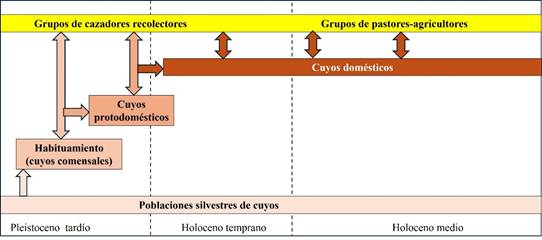
Figura 6
Proceso de domesticación del cuyo iniciada desde el Pleistoceno, gracias a su habilidad para aprovechar los campamentos humanos (elaborada por Raúl Valadez)
(elaborada por Raúl Valadez)
Figura 6. Proceso de domesticación del cuyo iniciada desde el Pleistoceno, gracias a su habilidad para aprovechar los campamentos humanos (elaborada por Raúl Valadez)
El guanaco (Lama glama guanicoe)
El segundo animal americano involucrado en la domesticación fue el guanaco (Valadez, 2023: 284-291), derivándose de este proceso la llama (Lama glama glama).
Las evidencias al respecto dependen por completo de las características físicas de cada tipo y la clase de evidencia arqueozoológica disponible. Los guanacos son de talla menor (un metro de alzada) y su pelaje varía del rojo claro al blanco, siendo “tipo lanilla”, es decir, no grueso y más bien heterogéneo; mientras que en la llama se presentan diversas tonalidades, es más grueso y homogéneo. A nivel osteológico existen diversos detalles en la forma del cráneo y morfología de las falanges que permiten, de forma general, diferenciar entre guanacos, llamas o camélidos con caracteres mezclados, los cuales podrían haber correspondido a ejemplares protodomésticos (Valadez, 2023: 290-291).
La información disponible muestra un proceso de adaptación largo, pasando por las diferentes fases desde el Pleistoceno superior (Figura 4). Todo indica que la relación dominante entre guanaco y humanos fue siempre de cacería; sin embargo, la constante búsqueda de alimento, agua y refugio promovió que en el páramo andino unos y otros mantuvieran un estrecho contacto.
Un sitio interesante es la Quebrada de Humahuaca en la provincia de Jujuy en el noroeste de Argentina (Figura 7), donde se estudió un refugio al pie de una ladera rocosa (Elkin y Rosenfeld, 2001: 49). Su temporalidad fue muy diversa, desde inicios del Holoceno hasta tiempos coloniales. En niveles de una temporalidad sobre los ocho mil años o poco más, se reconoció una falange de Lama glama glama, es decir, una llama como tal, al interior de un espacio habitacional y directamente ligado con actividades de procesamiento y consumo de alimento. Esta temporalidad y el tipo de hallazgo quizá refiera a un momento en que ya existían poblaciones de guanacos lo bastante modificados para que se manifestaran dichos cambios a nivel anatómico; aunque lo escaso de la muestra, su temporalidad y su aislamiento respecto de otros casos equivalentes quizá indique la presencia de poblaciones protodomésticas, es decir, bastante adaptadas al ámbito humano como para que se manifestaran las adaptaciones endócrinas a nivel morfológico, pero sin que el hombre tuviera aún una participación activa.
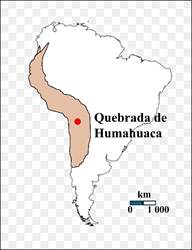
Figura 7.
Ubicación del sitio de Humahuaca, donde se descubrió una falange de llama de ocho mil años de antigüedad. Este registro evidencia la posible existencia de llamas protodomésticas para este momento
Figura 7. Ubicación del sitio de Humahuaca, donde se descubrió una falange de llama de ocho mil años de antigüedad. Este registro evidencia la posible existencia de llamas protodomésticas para este momento
En el contexto arqueológico, los restos de guanaco aparecen desde el Pleistoceno y en todo el Holoceno. Trátese de huesos, dientes o pelo, no hay duda que su presencia es una constante, aunque los números varíen en función del lugar y el momento.
Dentro de los registros más antiguos se han encontrado ejemplares “tipo guanaco”, pero de características peculiares (Mengoni y Yacobaccio, 2006: 239); por un lado, se tienen los que son de talla más grande, fenómeno que también se observa en algunas localidades más tardías y que, por otro lado, ha derivado a la opción de que correspondan a organismos en proceso de domesticación. Caso equivalente es el hallazgo de pelo cuyas características son más semejantes a los de la llama, aunque se ve más probable que se trate de variaciones individuales.
En el periodo entre el fin del Pleistoceno y los ocho o siete mil años hasta ahora, la relación hombre-guanaco está dominada por la cacería. Dentro de dicho lapso se observa una intensificación en la presencia de restos de este camélido, sobre todo a partir de los ocho mil quinientos años antes del presente en adelante. Entre este momento y los seis mil años es frecuente la referencia de “camélidos grandes o que tienden al patrón llama”, presumiblemente animales protodomésticos (Figura 8). Cronológicamente estos cambios morfológicos poco a poco van también acompañados de mayores abundancias de las “protollamas” en los contextos arqueológicos en comparación, por ejemplo, con los venados, lo cual demuestra una gran intensidad en su empleo, sin duda por la copiosa cantidad de los rebaños alrededor de los espacios humanos, todo lo cual habría tenido lugar sobre los siete mil años a. p.
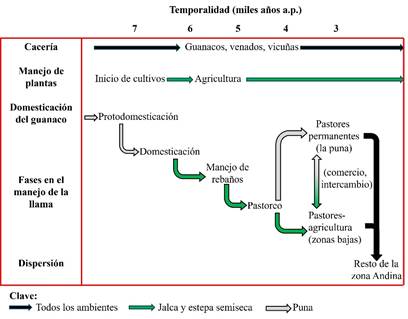
Figura 8
Proceso de evolución de la domesticación del guanaco hacia la llama y hasta el pastoreo (elaborado por Raúl Valadez)
Figura 8. Proceso de evolución de la domesticación del guanaco hacia la llama y hasta el pastoreo (elaborado por Raúl Valadez)
A partir de este momento las evidencias a favor de la aparición de las llamas se hacen más profusas: morfología, presencia de rebaños, coloraciones propias de este animal, diversidad de edades, restos de corrales, todo lo cual se consolida sobre los seis mil años a. p. Acaso mucho del proceso se aceleró conforme los grupos humanos fueron aprendiendo el manejo de plantas pues, conforme esta actividad se consolidó, se aseguró el abasto de alimento. No obstante debe hacerse notar que en gran parte del mundo andino la actividad de pastoreo se desarrolló de forma independiente a lo agrícola, convirtiéndose en una forma de interacción que ha perdurado hasta el presente.
El guajolote (Meleagris gallopavo gallopavo)
El tercer caso, en orden cronológico, corresponde al guajolote, cuya evolución tuvo lugar en territorio mexicano.
En este caso las evidencias son menos claras que en los anteriores, pero suficientes para reconstruir el proceso (Figura 4). El punto de inicio es la consideración de que esta ave tiene como hábitat primario los bosques templados (Figura 9), mas es igualmente hábil para introducirse en biomas como pastizales o bosques tropicales caducifolios. En territorio mexicano, sobre todo en la región central y sur, estos ecosistemas coinciden en diversos sitios, lo cual ofrece la opción de que los cambios ocurridos al inicio del Holoceno hayan derivado en poblaciones cuya sobrevivencia dependía de sus habilidades adaptativas, incluyendo las que involucraban el espacio antropógeno. (Medina et al., 2020: 23-26; Valadez, 2020: 192-194).
Otro segundo aspecto relevante es que los estudios de ADN han demostrado que todo guajolote doméstico existente en el presente es descendiente de la subespecie Meleagris gallopavo gallopavo, cuyo rango de distribución abarcaba, justamente, el centro de México (Medina et al., 2020: 18-21) (Figura 9).
La tercer clave a considerar es la muestra arqueozoológica (Figura 9). Los restos más arcaicos tienen una antigüedad que va desde los 2,500 a los 3,200 años a. p., ciertamente se trata de hallazgos tardíos (en la perspectiva del Holoceno), pero corresponden a ejemplares que ya son de condición doméstica, conclusión derivada de la morfología ósea y de su presencia en contextos relacionados con comunidades agrícolas. Estos casos conciernen también a localidades del centro de México (Medina et al., 2020: 34-37; Valadez 2023: 194-195).
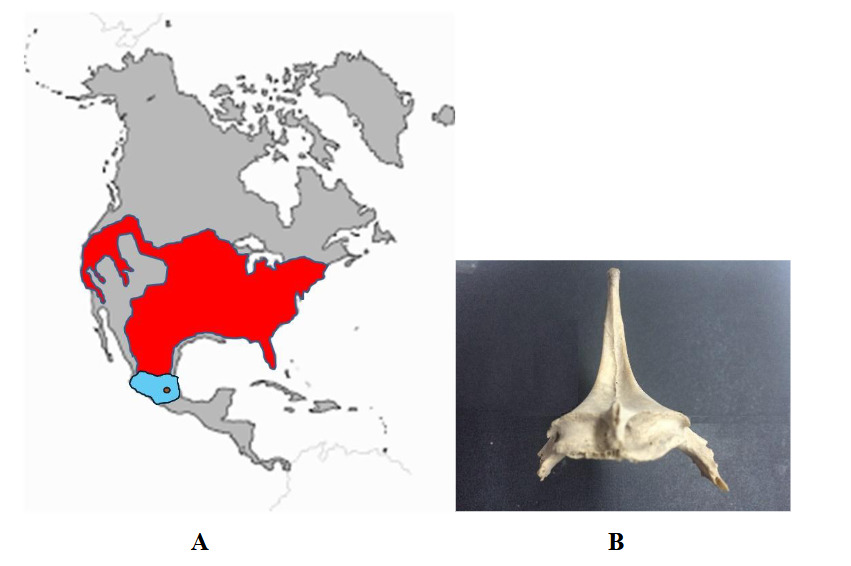
Figura 9
(A) Mapa de distribución de la especie Meleagris gallopavo (zona coloreada de rojo y azul), región donde habitó la subespecie M. gallopavo gallopavo (espacio azul) y sitios donde se han encontrado los más antiguos restos (círculo). (B) Vista anterior de una quilla de guajolote descubierta en el sitio de Huixtoco, cuya antigüedad fue de 2,500 años a. p. y cuya morfología es, indudablemente, la propia de un ejemplar doméstico (Medina et al., 2020: 34-37; elaborado por Raúl Valadez)
Figura 9. (A) Mapa de distribución de la especie Meleagris gallopavo (zona coloreada de rojo y azul), región donde habitó la subespecie M. gallopavo gallopavo (espacio azul) y sitios donde se han encontrado los más antiguos restos (círculo). (B) Vista anterior de una quilla de guajolote descubierta en el sitio de Huixtoco, cuya antigüedad fue de 2,500 años a. p. y cuya morfología es, indudablemente, la propia de un ejemplar doméstico (Medina et al., 2020: 34-37; elaborado por Raúl Valadez)
Respecto del factor humano, a partir del Holoceno temprano el modo de vida de las bandas va cambiando en la mayor parte del continente hacia un sistema de macrobandas estacionales que permanecían tiempos mayores en zonas donde los recursos y el clima eran más favorables y cuando eso cambiaba se disgregaban. Este patrón por temporadas provocó que ya desde los ocho mil años a. p. en adelante se tengan evidencias de plantas que son aprovechadas y seleccionadas por estos grupos; un par de miles de años después, su cultivo es una realidad (Figura 4).
El guajolote silvestre es un animal gregario que pasa casi todo el tiempo en tierra, anida al interior de macollos y requiere de fuentes de agua permanentes. Se alimenta de pequeños animales como insectos, semillas y restos vegetales diversos (Valadez, 2023: 192). Esta tendencia debió haber auxiliado en su habituamiento con macrobandas en lugares donde el clima variaba de acuerdo con la época del año y mientras la propensión en el Holoceno temprano se encaminaba hacia temperaturas más altas y a menor precipitación, o al menos a un acentuamiento de la estacionalidad.
La biología de esta ave y su relación con las comunidades humanas y espacios de cultivo han demostrado que su paulatina adaptación a lo antropógeno estuvo íntimamente ligada al sedentarismo y desarrollo del agroecosistema de la milpa (Valadez, 2023: 202-204), lo cual significa que su habituamiento tuvo lugar simultáneamente a los cambios en el esquema de vida de estas comunidades. El hecho de que ya hace tres mil años se reconozca la presencia de poblaciones domésticas y completamente integradas a lo humano significa que este último nivel pudo haber empezado un par de miles de años antes y que la etapa de protodomesticación tuvo lugar entre los cinco y siete mil años a. p., época en la que se tienen las más tempranas evidencias del cultivo de plantas como el maíz, la calabaza y el quenopodio (McClung y Zurita, 2014: 274).
La vicuña (Lama glama vicuña)
Viajando de nuevo al otro lado del continente y también como un caso ya involucrado con el desarrollo de la agricultura tenemos a la alpaca (Lama glama pacos) (Valadez, 2023: 292-296). Esta forma doméstica se derivó de la vicuña, cuyo proceso en este sentido fue parecido al del guanaco, pero más tardío. Los hallazgos más antiguos son de hace unos seis mil años en los andes peruanos, ya en espacios donde la agricultura era una realidad. Se considera que el proceso inició en el Holoceno temprano, hace unos 8,000 años (Figura 4).
El ancestro silvestre de la alpaca es la vicuña (Lama glama vicuña). Su distribución abarcaba zonas altas y húmedas de los Andes, desde el norte de Perú hasta el norte de Chile. Su talla es menor a la del guanaco y similar a la de su derivado doméstico. Tiene una piel cubierta de lana fina y densa, pero de longitud corta; mientras que la de la alpaca es de un grosor medio pero mucho más larga.
Las evidencias del paso de poblaciones de vicuñas hacia el tipo alpaca son fragmentadas y limitadas (Valadez, 2023: 294-296), aunque podemos considerar que todo ello se inició al final del Holoceno temprano, hace unos ocho mil años (Figura 4). Los estudios de ADN vinculan a las poblaciones de vicuñas del centro de los Andes peruanos con el proceso, por lo que se considera a esta región como su lugar de origen (Marín et al., 2007: 137-138) (Figura 10).
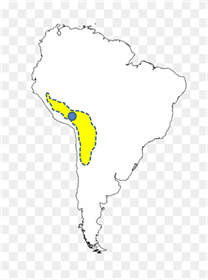
Figura 10
Distribución natural de la vicuña y posible lugar de origen de la alpaca (círculo azul), conclusión derivada de registros de dientes y de estudios del ADN (Valadez, 2023: 2
Figura 10. Distribución natural de la vicuña y posible lugar de origen de la alpaca (círculo azul), conclusión derivada de registros de dientes y de estudios del ADN (Valadez, 2023: 296)
A nivel arqueozoológico, el más importante elemento diagnóstico para reconocer a la alpaca son los incisivos, que poseen forma espatulada con esmalte en la cara labial; a diferencia de los de las vicuñas, que son de forma rectangular con el mismo esquema de esmalte, o los de llamas y guanacos que tienen esmalte en ambas caras (Valadez, 2021: 83). De acuerdo con esto, piezas dentales “tipo alpaca” se han reconocido en sitios del centro de los Andes peruanos cuya antigüedad se ubica sobre los seis mil años a. p. (Mengoni y Yacobaccio, 2006: 240; Valadez, 2023: 295), siendo tres mil años después cuando se dispone de evidencia clara sobre su presencia.
De acuerdo con esto, y un poco semejando el caso del guajolote, es la conjunción de estos diversos elementos lo que permite establecer un probable esquema de desarrollo de la domesticación de la vicuña. La muestra de dientes, indudable pero aislada, habla de un momento en el que quizá ya existían poblaciones en la fase de protodomesticación ligadas al espacio agrícola (Figura 4). Dado que en esa época (Holoceno medio) la llama es una realidad, así como las comunidades agrícolas y pastoras, quizá el control de las poblaciones protodomésticas existentes desde los 6,000 años a. p., ya con su incipiente cubierta de lana, promoviera el interés para impulsar el proceso, derivándose de ello la aparición de las alpacas hace tres mil años o incluso antes.
Discusión
A través de los casos presentados hemos visto cómo es que tuvo lugar la aparición de fauna doméstica al inicio del Pleistoceno y que relación guarda con cambios ambientales y las comunidades humanas.
Tal y como se indicó al inicio, este fenómeno para esa época estuvo fuertemente ligado a las modificaciones ecológicas que tuvieron lugar en América y otras regiones del mundo, principalmente Medio Oriente, promoviendo formas de interacción que en un principio tenían como móvil los beneficios que estos animales obtenían, sin que al ser humano le afectara en sentido positivo o negativo. Este esquema, al que biológicamente se le denomina “comensalismo”, se mantuvo mientras tuvo lugar el habituamiento y la protodomesticación; fue sólo hasta que las personas tomaron conciencia de las características de esa fauna, permanentemente ligada a su espacio, que tomaron paulatinamente el control de dichas poblaciones, dando paso así a la domesticación como tal, la cual es una relación de simbiosis en tanto que unos y otros se nutren a través de ésta (Valadez, 2021: 24-29).
Si consideramos la situación de que los más antiguos animales domésticos surgieron en más de un lugar en el mundo: Medio Oriente, África, Asia central, Asia sudoriental, América del Norte y América del sur, todos fuertemente ligados a los cambios ambientales mencionados, entonces debemos reconocer este proceso como estrategia de sobrevivencia y no como manifestaciones de la inteligencia humana. Por qué abarcó sólo a ciertas especies, es simplemente la condición de los seres vivos, cuya capacidad para adaptarse y subsistir depende de innumerables factores.
Centrándonos más en nuestro tema, debemos enfatizar la relevancia de darle un espacio de investigación óptimo, puesto que fácilmente se cae en el error de verlo como algo poco importante dentro del desarrollo de las culturas americanas. No obstante que en algunos casos como el de los camélidos existe una enorme cantidad de artículos sobre su presencia, en realidad es muy poco lo que se trabaja para entender la forma en como inició el proceso, dónde y cuándo. En otros casos, como el cuyo, sin duda es muy interesante saber la manera en como este roedor se fue integrando a los grupos humanos, desde cuándo y cómo se fue creando la simbiosis para con el hombre, que sin duda lo vio siempre como una presa fácil y siempre disponible, e, incluso así, las gracias para el animal fueron bastante grandes, de modo que su condición doméstica se hizo realidad.
Respecto del guajolote, el gran problema ha sido siempre presuponer que su presencia fue continua, constante, perdida en el pasado, de forma que sencillamente se da por hecho que no tiene sentido su estudio ni su origen, pues a fin de cuentas se le trata como un animal doméstico presente en todas partes y desde siempre. Lo que se presentó en páginas anteriores demuestra que este pensamiento es indudablemente falso y que innegablemente falta mucho por saber respecto de su origen.
Reflexiones finales
Como se observa en la Figura 4, los casos aquí presentados no son, ni por mucho, lo que en esta época se conoce como fauna doméstica americana. Antes bien, no sería nada improbable que en casos como las abejas, al paso del tiempo sea posible reconocer elementos que permitan retroceder el momento de su condición doméstica a, por ejemplo, la aparición de plantas domésticas y así considerar que se originaron en el Holoceno medio, hace seis o hasta siete mil años, y con ello, todos los beneficios asociados. De esta forma es importante no perder de vista este tema y, sobre todo, asignarle un papel individual al momento de estudiar la fauna del Pleistoceno superior y Holoceno inferior y medio, ya que ahí se encuentran las evidencias sobre el origen de esta fauna y de todo lo que involucra su estudio y su impacto en las sociedades humanas de América.
Referencias
www.mma.gob.cl
Altamirano, Alfredo (1986). “La importancia del cuy: un estudio preliminar, Perú”. Serie investigaciones No 8.Lima: Facultad de Ciencias Sociales, Universidad Nacional Mayor de San Marcos.
Ardelean, Ciprian; Becerra, Lorena; Pedersen, Mikkel; Schwenninger, Jean-Luc; Oviatt, Charles; Macías, Juan; Arroyo, Joaquín; Sikora, Martín; Ocampo, Yam Zul; Rubio, Igor; Watling, Jennifer; de Medeiros, Vanda; De Oliveira, Paulo; Barba, Luis; Ortiz, Agustín; Blancas, Jorge; Rivera, Irán; Solís, Corina; Rodríguez, María; Gandy, Devlin; Navarro, Zamara; De La Rosa, Jesús; Huerta, Vladimir; Marroquín, Marco; Martínez, Martin; López, Alejandro; Higham, Thomas, y Willerslev, Eske (2020). “Evidence of Human Occupation in Mexico around the Last Glacial Maximum”. Nature. Consultado el 15 de agosto de 2020 en https://doi.org/10.1038/s41586-020-2509-0
Elkin, Dolores y Rosenfeld, Silvana (2001). “Análisis faunístico de Pintoscayoc 1 (Jujuy)”, pp. 29-64. En Mengoni, Guillermo; Olivera, Daniel y Yacobaccio, Hugo (eds.) El uso de los camélidos a través del tiempo. Buenos Aires, Argentina: Grupo Zooarqueología de Camélidos, International Council for Archaeozoology, Ediciones del Tridente.
Hall, Raymond (1981). The Mammals of North America. Vol II. Estados Unidos: The Ronald Press Company.
Hesse, Brian (1984). “Archaic Exploitation of Small Mammals and Birds in Northern Chile”. Estudios Atacameños 7, pp. 37-51. DOI: 10.22199/S07181043.1984.0007.00005.
Ijzereef, Gerard (1978). “Faunal Remains from the El Abra Rock Shelters (Colombia)”. Palaeogeography. Palaeoclimatology. Palaeoecology 25(1-2), pp.163–177.
Lord, Edana; Collins, Catherine; de France, Susan; LeFebvre, Michelle; Pigière, Fabienne; Eeckhout, Peter; Erauw, Celine; Fitzpatrick, Scott; Healy, Paul; Martínez, M. Fernanda; García, J.; Ramos, Elizabeth; Delgado, Miguel; Sánchez, Ancira; Peña, Germán; Toyne, J.; Dahlstedt, Un; Moore, Katherine; Laguer, C.; Zori, Collen, y Matisoo, Lisa (2020). “Ancient DNA of Guinea Pigs (Cavia spp). Indicates a Probable New Center of Domestication and Pathways of Global Distribution”. Scientific Reports 10. 8901 en https://doi.org/10.1038/s41598-020-65784-6
McClung, Emily y Zurita, Judith (2014). “Las primeras sociedades sedentarias”, pp. 255-296. En Manzanilla, Linda R. y López, Leonardo (coords.). Historia antigua de México, vol I. México: Consejo Nacional para la Cultura y las Artes, Instituto Nacional de Antropología e Historia, Miguel Ángel Porrúa, Universidad Nacional Autónoma de México.
Medina, Andrés; Valadez, Raúl; Pérez, Gilberto, y Rodríguez, Bernardo (2020). Huexolotl, pasado y presente en México. México: Universidad Nacional Autónoma de México.
Mengoni, Guillermo y Yacobaccio, Hugo (2006). “The Domestication of South American Camelids. A View from the South-Central Andes”, pp. 228-244, capitulo 16. En Zeder, Melinda; Daniel, Bradley; Emshwiller, Eve, y Smith, Bruce (eds.). Documenting Domestication. New Genetic and Archaeological Paradigms. Estados Unidos-Inglaterra: University of California Press.
Rofes, Juan y Wheeler, Jane (2003). “Sacrificio de cuyes en los Andes: el caso de El Yaral y una revisión biológica, arqueológica y etnográfica de la especie Cavia porcellus”. Archaeofauna 12, pp. 29-45.
Valadez, Raúl (2021). Los animales domésticos. Su estudio, su origen, su historia. Tomo I El fenómeno de la domesticación animal y su estudio. México: Universidad Nacional Autónoma de México.
(2023). Los animales domésticos. Su estudio, su origen, su historia. Tomo II La domesticación animal en el mundo antiguo. México: Universidad Nacional Autónoma de México.
Valadez, Raúl y Pérez, Gilberto (en prensa). “Registros pleistocénicos del Rancho Córdova y sus implicaciones con el registro fósil de perros en América”. En Pérez, Gilberto (ed.). Rancho Córdova, Cedral, San Luis Potosí: exploración y estudio de sus materiales. México: Facultad de Ciencias Sociales y Humanidades, Universidad Autónoma de San Luis Potosí.
Notas
Información adicional
redalyc-journal-id: 8006