ARTÍCULO ORIGINAL
Análisis espacio-temporal de las condiciones biofísicas y ecológicasde Triatoma dimidiata (Hemiptera: Reduviidae: Triatominae)en la region nororiental de los Andes de Colombia
Spatio-temporal analysis of the biophysical and ecological conditions of Triatoma dimidiata (Hemiptera: Reduviidae: Triatominae) in the northeast region of Colombia
Análisis espacio-temporal de las condiciones biofísicas y ecológicasde Triatoma dimidiata (Hemiptera: Reduviidae: Triatominae)en la region nororiental de los Andes de Colombia
Biomédica, vol. 37, núm. 2, 2017
Instituto Nacional de Salud
Recepción: 25 Agosto 2016
Aprobación: 26 Julio 2017
Resumen
Introducción. : Ante el desconocimiento del contexto espacio-temporal de las condiciones biofísicas(hidrometereológicas y de densidad de la cobertura vegetal) de las zonas con presencia deTriatomadimidiata en Santander y Boyacá, es necesario dilucidar los patrones asociados con estas variablespara determinar su distribución y control.
Objetivo. : Hacer el análisis espacio-temporal de las variables biofísicas relacionadas con la distribución de Triatoma dimidiata de los departamentos de Santander y Boyacá en la región nororiental de Colombia.
Materiales y métodos. : Se utilizaron las bases de datos de los registros de presencia deT. dimidiatay defactores hidrometereológicos del Special Report on Emissions Scenarios (SRES) del IntergovernmentalPanel on Climate Change,IPCC. Se estudiaron las variables de temperatura, humedad relativa, pluviosidad y densidad de la cobertura vegetal a nivel regional y local, se hizo el análisis espacial y elgeoestadístico, así como el análisis estadístico descriptivo y temporal de las series de Fourier.
Resultados. : En las áreas con mayor presencia de T. dimidiata,las temperaturas a dos metros delsuelo y en suelo cubierto oscilaron entre 14,5 y 18,8 °C, y la temperatura ambiente fue de 30 a 32 °C. La densidad de la cobertura vegetal y la pluviosidad en las áreas de mayor presencia de T. dimidiataexhibieron patrones de picos anuales y bienales. Los valores de la humedad relativa fluctuaron entre66,8 y 85,1 %.
Conclusiones. : Las temperaturas a nivel de superficie y a dos metros del suelo fueron las variablesdeterminantes de la distribución espacio-temporal de T. dimidiata. La elevada humedad relativaincentivó la búsqueda de refugios e incrementó la distribución geográfica en los picos anual y bienalde pluviosidad a nivel regional. Las condiciones ecológicas y antrópicas sugieren que T. dimidiata esuna especie de gran resiliencia.
Palabras clave: Triatominae+ enfermedad de Chagas+ ecología+ clima+ análisis espacial+ Colombia.
Abstract
Introduction: Due to the lack of information regarding biophysical and spatio-temporal conditions (hydrometheorologic and vegetal coverage density) in areas with Triatoma dimidiata in the Colombiandepartments of Santander and Boyacá, there is a need to elucidate the association patterns of thesevariables to determine the distribution and control of this species.
Objective: To make a spatio-temporal analysis of biophysical variables related to the distribution of T.dimidiate observed in the northeast region of Colombia.
Materials and methods: We used the Intergovernmental Panel on Climate Change Special Report onEmissions Scenarios (IPCC SRES) data bases registering vector presence and hydrometheorologicdata. We studied the variables of environmental temperature, relative humidity, rainfall and vegetalcoverage density at regional and local levels, and we conducted spatial geostatistic, descriptive statistical and Fourier temporal series analyses.
Results: Temperatures two meters above the ground and on covered surface ranged from 14,5°C to18,8°C in the areas with the higher density of T. dimidiata. The environmental temperature fluctuatedbetween 30 and 32°C. Vegetal coverage density and rainfall showed patterns of annual and biannualpeaks. Relative humidity values fluctuated from 66,8 to 85,1%.
Conclusions: Surface temperature and soil coverage were the variables that better explained the lifecycle of T. dimidiata in the area. High relative humidity promoted the seek of shelters and an increaseof the geographic distribution in the annual and biannual peaks of regional rainfall. The ecologic andanthropic conditions suggest that T. dimidiata is a highly resilient species.
Key words: esENTriatominae, Chagas disease, ecology, climate, spatial analysis, Colombia.
En Colombia, varias especies de triatominos sonvectores deTrypanosoma cruzi (1,2), entre ellos,Triatoma dimidiata (3). Esta especie, originaria delsuroriente de México y nororiente de Guatemala(4), se extiende desde México hasta Ecuador y elnoreste de Perú (5). Se sabe que T. dimidiata tieneuna gran variabilidad intraespecífica (3,6-10) y queocupa ambientes tanto selváticos como domésticosy peridomésticos (3,5,7,8).
Esta especie, típica de hábitats terrestres (11), se adapta a diferentes ecótopos (cuevas, rocas,oquedades, nidos de vertebrados, etc.) en elambiente selvático (8,11-14), incluidas las palmasde la especie Attalea butyracea en el Caribecolombiano y en la Sierra Nevada de Santa Marta(15). Además, es una especie capaz de sobreviviren diferentes rangos altitudinales (12), con laconsecuente capacidad de invasión y dispersiónespacial y estacional (8). Su domiciliación se vefavorecida por las condiciones ambientales (2),principalmente en la región centro-oriental del país,en los departamentos de Boyacá y Santander(12,16). Sin embargo, es poco lo que se sabesobre las condiciones biofísicas que favorecen suproliferación (12,17).
Además del estudio de su biología y ecoepidemiología,ladistribucióngeográficadeotrasespeciesdetriatominos (por ejemplo, T. infestans, T.patagonica, T. platensis y Rhodnius spp.), seha relacionado con variables biofísicas como la temperatura, la humedad, la precipitación y la elevacion (18). En este sentido, se ha demostradoque, en la península de Yucatán, T. dimidiata esmás abundante en los domicilios y que la tasa deinfección por T. cruzi es mayor en la época de calorentre abril y junio (19).
Asimismo, se ha comprobado que los vectoresdel género Triatoma localizados en zonas geográficascon altas temperaturas pueden tener másgeneraciones por año (18) que los de las zonas más templadas (20), y que la mayor diversidad deespecies de triatominos se localiza en biomas conestaciones seca y húmeda definidas y con largastemporadas de temperaturas altas (21-24), lo cualresulta en grandes densidades poblacionales yejemplares con una mayor capacidad de dispersiónhacia hábitats artificiales (22,24).
En el presente estudio se tuvieron en cuenta lascondiciones climáticas publicadas por el IntergovernmentalPanel on Climate Change (IPCC),en especial las proyecciones del Special Reporton Emissions Scenarios (SRES) de 2013-2014(25) relativas a la temperatura ambiente (Tamb), latemperatura a dos metros del suelo (T2m) y a nivelde superficie (Tms).
La temperatura ambiente o del aire se define comola medida de calor sensible que se almacena enél (26) en un momento determinado, incluida suevolución espacio-temporal en un área dada. Estaes la base para la diferenciación de los tipos climáticos,en especial en la clasificación de Köppen(27), cuya sencilla división permite clasificar losclimas como macrotérmicos, con temperaturas porencima de 18 °C a lo largo de todo el año; comomesotérmicos o templados, con temperaturas entre-3 y 18 °C; como más cálidos, con una temperaturapor encima de los de 10 °C, y como microtérmicoso fríos, con temperaturas menores de 10 °C entodo el año.
En los últimos años ha aumentado el interés porlos estudios de ecoepidemiología y epidemiologíaespacial, dada la facilidad que ofrecen los sistemasde información geográfica (SIG), el acceso a losdatos provenientes de los sensores remotos (28-36)y el desarrollo de sistemas computacionales quepermiten la aplicación de algoritmos complejos enun tiempo relativamente reducido mediante técnicasde modelado, discriminatorias, descriptivas y mixtas(37,38), y en diversas escalas espacio-temporales(39), lo cual ofrece una mejor perspectiva y unaaproximación más realista a los factores de riesgode la transmisión de enfermedades infecciosas (40).
En Colombia, se ha aplicado este tipo de herramientas,incluidas las técnicas geoestadísticas y lassimulaciones de focos potenciales de infestación, al estudio de T. dimidiata (12), y los datos obtenidoshan servido para adoptar otros enfoquesmetodológicos (Parra-Henao G, Badel-MogollónJE. Uso de SIG, SR y análisis espaciales para lageneración de modelos predictores de distribuciónde triatominos (Hemiptera: Reduviidae), vectores deenfermedad de Chagas en Colombia. Biomédica.2015;35 (Supl.4):28-30. Memorias, XVI CongresoColombiano de Parasitología y Medicina Tropical).En este estudio de T. dimidiata en Colombia (12),se analizaron los factores ambientales determinantesde la distribución de la especie mediante unmodelo de regresión logística de la probabilidad depresencia del vector desarrollado con el algoritmoMaxEnt. En él se encontró una asociación significativaentre la presencia del vector, los valoresmínimos de la temperatura de la superficie terrestre,la mayor densidad de la cobertura vegetal en elárea de estudio y la elevación del terreno, lo cualdio lugar a nuevas preguntas sobre las condicionesambientales favorables a las poblaciones deT. dimidiata, en particular, y de los triatominos en general.
El algoritmo MaxEnt funciona como una caja negra,similar a los modelos cuyo método se basa enenfoques bayesianos y de redes neurales, a losmodelos aditivos generalizados, y a los linealesgeneralizados (41). Pero aunque dicha aproximaciónpermiteentendermejorelconjuntoyexplicarclaramentelapresenciadelvectorenlasáreasderecoleccion,su naturaleza como modelo del tipode "caja negra" dificulta el conocimiento real de losvalores y rangos de las variables que mejor explicanla presencia del vector.
Teniendo en cuenta esta consideración, una vezconocidos los mayores focos potenciales de infestación,elpasosiguientehasidoexplicarlapresenciadeT.dimidiata,peroconbaseenlosvaloresobservadosenlasvariablesbiofísicasconsideradascomode mayor relevancia y en otras que no seincluyeron en los modelos iniciales, pero que puedenser importantes en la distribución del vector.
Además de explicar la distribución espacial de T.dimidiata en Colombia, también se procuró dilucidaralgunos detalles de las fluctuación temporalde las poblaciones. Para ello se analizaron lasseries temporales de algunas variables físicas yecológicas básicas utilizando las transformadasde Fourier (42).
El análisis armónico, o análisis de Fourier, seutiliza para el procesamiento digital de imágenes,principalmente en física e ingeniería, aunque hoy se aplica en muchos campos, desde la predicciónde la tendencia de las llamadas en un centropolicial (Lewis B, Herbert R, Bell R. The applicationof Fourier analysis to forecasting the inbound calltime series of a call centre. Proceedings of theInternational Congress on Modeling and SimulationMODSIM03; Townsville, Australia. 2003. p.1281-6), hasta el análisis en imágenes de las seriestemporales del índice de vegetación (43) y dediversos fenómenos ambientales (44).
En síntesis, el objetivo de este trabajo fue determinarlas condiciones ecológicas en el área deinfluencia de los focos potenciales de infestaciónde T. dimidiata en la región andina con base enun análisis espacio-temporal, y su relación condiversas variables biofísicas y con la coberturavegetal mediante el modelado geoestadístico, lasimulación condicional y el análisis de Fourier.
Materiales y metodos
Área de estudio
Se elaboró una base de datos en la cual se registróla presencia de T. dimidiata en viviendas muestreadasenlosmunicipiosdeSoatáyTipacoqueeneldepartamentode Boyacá, y Capitanejo, Enciso,Macaravita, Molagavita, San Vicente de Chucurí ySan José, en el departamento de Santander.
Con base en la información geoestadística delas áreas con mayor presencia de T. dimidiataen Colombia, se escogió una zona del nororientede la Región Andina localizada entre los 6,2° y7,0° de latitud norte, y los 72,2° y 73,0°de longitudoeste, específicamente entre las subregionesnaturales de las montañas santandereanas, elcañón de los ríos Chicamocha y Suárez, el sur delmacizo de Santurbán, el nororiente de las laderasde la Cordillera Oriental y el norte de la planiciecundiboyacense (figura 1a). En estas coordenadasse localizan los municipios pertenecientes a losdepartamentos de Boyacá (Tipacoque y Soatá)y de Santander (Capitanejo, Enciso, Macaravita,Molagavita, San José y San Vicente de Chucurí), endonde se ubicaron los ocho puntos de recolecciónde triatominos.
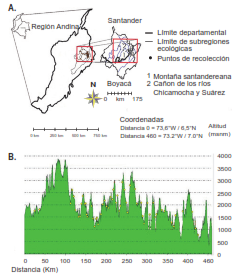
Figura 1
A. Región Andina de Colombia. En el recuadro se ve el área de recolección. B. Perfil altitudinal de los puntos de recolección de T. dimidiata en los ecótopos de la montaña santandereana (a) y el cañón del Chicamocha (b). En el recuadro se destaca con puntos amarillos la altitud de los puntos de recolección de T. dimidiata, incluidos el punto más elevado y el menos elevado, según la ruta mostrada en el mapa con el trazo sólido de color amarillo. Los trazos sólidos de color negro representan los límites de los ecótopos. Mapa elaborado a partir del modelo digital de elevación (MDE) del SRTM Worldwide Elevation Data, con 3 arcosegundos de resolución
La altitud de estos puntos fluctuó entre los 755 my los 2.938 m (figura 1b), lo que supone una granvariación en los valores de la temperatura ambientey de la superficial, entre otros, pues abarcan todoslos pisos térmicos y condiciones climáticas, desdeel clima templado seco del cañón del Chicamochahasta los climas templados y fríos.
Modelos de temperatura y humedad relativa media del ambiente en las áreas de recolección
Para el modelado descriptivo de las variables bio-climáticas a nivel macro, relevantes para explicar las condiciones ecológicas que rigen la distribución deT. dimidiata en el área de estudio, se utilizaron losdatos del Modelo Climático Regional Eta-CPTECdel Instituto Nacional de Pesquisas Espaciais(INPE) de Brasil, los cuales incluyen proyeccionessobre el cambio climático en Suramérica con unaresolución espacial de 40 km para los períodos de1961 a 1990 y de 2010 a 2100 en las condicionesde medias de emisiones SRES A1B (45) del modeloglobal Hadley Centre Coupled Model, versión 3(HadCM3), ofrecidas por el Metoffice Hadley Centre(MOHC) del Reino Unido en el escenario A1B, esdecir, crecimiento económico y poblacional rápidoen un mundo globalizado (acentuada interacción yconvergencia entre países).
El modelo HadCM3 es del tipo atmósfera-océano,de resolución atmosférica vertical terrestre de 19niveles y 14 bandas espectrales, y no necesitaningún tipo de ajuste de flujo para producir buenassimulaciones; el modelo funciona con base en elconcepto de Perturbed Physics Ensembles (PPE).En el artículo de Gordon, et al. (46), puede hallarseuna descripción detallada del modelo.
Se tuvieron en cuenta los datos del subconjunto decontrol de las situaciones en que la temperatura esun parámetro resultante de la media de algunas desus variables: temperatura ambiente (Tamb), temperaturaadosmetrosdelasuperficiedelsuelo(T2m),temperaturaaniveldelasuperficiedelsuelo(Tms)ytemperatura en el suelo cubierto (Tsc). La últimavariable estudiada en el marco del modelo HadCM3fue la humedad relativa media del ambiente.
Pluviosidad y cobertura vegetal
Además del modelo HadCM3, también se consideróla pluviosidad en el área de los puntos de recolección.Losvaloresdelapluviosidadcorrespondieronalosdatosdelaprecipitaciónmensualacumuladaregistradospor la Tropical Rainforest MeasuringMission (TRMM), producto 3B43V6, con resoluciónespacial de 5 km, disponible para cualquier coordenadageográfica de Suramérica. Los datos dela TRMM son distribuidos por el Data and InformationServicesCenter(DISC)delDistributedActiveArchiveSystem(DACC),atravésdeGoddardEarthSciences(GES) de la NASA.
Dichos datos se interpretaron conforme a losresultados obtenidos en los análisis estadísticosdescriptivos y de acuerdo con la geolocalizaciónde cada área de muestreo, y luego se compararoncon los obtenidos para los departamentos y seinterpretaron conforme a los rangos de valores reportados en la bibliografía correspondiente.
Como variable ecológica, se tuvo en cuenta la densidaddelacoberturavegetalsegúnlosresultadosdelaaplicacióndelaversión2delíndicemejoradodevegetación(EnhancedVegetationIndex2,EVI2),elcualseaplicautilizandolasiguientefórmula(47):

donde NIR corresponde a la banda infrarroja cercanadel espectro electromagnético y Red, a labanda roja del espectro.
La principal ventaja de este índice es que nose ve afectado por puntos de saturación, comopuede suceder con el clásico índice normalizadodiferencial de la vegetación (Normalized Difference Vegetation Index, NDVI), lo cual lo hace ideal parasu uso en áreas de vegetación muy densa, comosucede en los trópicos (47-49).
El cálculo del EVI2 se hace con imágenes MODISdel producto MOD13, colección 005, con resoluciónespacial de 250 m. Los datos MODIS son distribuidospor el Land Processes Distributed Active ArchiveCenter (LP-DAAC) de la US Geological Survey(USGS), del centro Earth Resources Observationand Science (EROS) (https://lpdaac.usgs.gov).
Los datos de este cálculo se obtuvieron delLaboratorio de Sensoriamento Remoto Aplicadoà Agricultura e Floresta (LAF), del INPE de Brasil(http://www.dsr.inpe.br/laf/). En diversos trabajosllevados a cabo en zonas tropicales, se ha sugeridoque las amplitudes en los armónicos son bajascuando se aplica el EVI2 en bosques primarios ysecundarios.
Todas las variables se evaluaron en los meses demayo (mes de muestreo en el departamento deBoyacá en el 2007) y de junio (mes de muestreoen el departamento de Santander en el mismoaño). Al no disponerse de series de datos del 2007en el modelo HadCM3, se recurrió a las seriesde los años 2010 a 2015, tanto para las áreas derecoleccion de T. dimidiata como para las regionesecológicas de los departamentos incluidos. Estasseries se validaron con las series de datos delHadCM3 para esos meses entre 1981 y 1990. Los datos de los productos MODIS y TRMM sevalidaron con las series de los años 2000 a 2006 y2008 a 2014, respectivamente.
Análisis armónico
Para el análisis de un conjunto discreto de datos,se aplica la transformada discreta de Fourier(TDF). En este caso, se empleó específicamentela transformada rápida de Fourier (FFT), que esun algoritmo de eficiencia computacional para calcularlaTDF,asícomoellenguajePython,porelfácilmanejodesuscomandosysuamplialibreríacientífica(SciPy).
Enla ventana de tiempo objeto de análisis, latransformada de Fourier arroja dos valores: unaamplitud que indica la intensidad de variación dela serie temporal asociada a la periodicidad del evento, y una fase que toma valores entre 0 y 2p para indicar el momento en el periodo en que se dala máxima variación.
Las series de Fourier responden a la siguienteecuación básica:
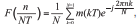
donde N es el número de muestras de la ventanaque se va a analizar, T es el período de muestreo,F es la frecuencia de muestreo, n es el índice de lafrecuencia cuyo valor se quiere obtener, y m (kT)indica la muestra tomada en el instante kT (muestra'késima') de la ventana.
Entre las coordenadas 6,2° y 7,4° N y 71,8° y 73,0° O en Boyacá y Santander, se obtuvieron datos mensualesde pluviosidad y cobertura vegetal desdeenero de 2001 hasta diciembre de 2012 en 49localidades diferentes. Se aplicó la transformadade Fourier a cada localidad y se graficó el resultadodel espectro de amplitud de la ondas en funcióndel tiempo, lo cual permite una mejor comprensiónde la periodicidad de dicho patrón en la señal y ladetección de algún posible patrón en las variablesbioclimáticas que pueda tener relación con el ciclode vida de T. dimidiata y facilite la adopción dedecisiones relacionadas con su control.
Resultados
Los datos mostraron que en Santander no serecolectaron ejemplares del vector en el domicilio,sino en ecótopos del peridomicilio o el extradomicilio,entantoqueenlosmunicipiosboyacencesseencontraronen el domicilio.
EnBoyacáhubo98reportesdetriatominosen28casasen el municipio de Soatá y 24 en sendascasas de Tipacoque. En San Vicente de Chucurí,Santander, se obtuvieron 17 reportes en cincocasas; en Macaravita, ocho en una casa; enMolagavita, nueve en una vivienda; en Enciso, seisen una vivienda; en Capitanejo, 50 en seis casas,y en San José, seis reportes en una vivienda. Losdatos de los municipios de Boyacá correspondieronal mes de mayo, y las de Santander, al mes dejunio de 2007.
Temperatura ambiente media
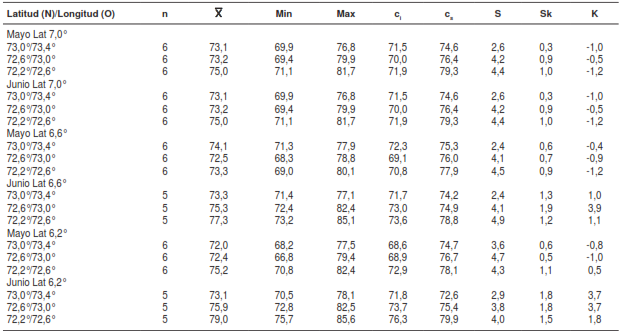
En el cuadro 1a, en la franja latitudinal más ecuatorialdelaslocalidadesdemuestreo(6,2°y6,6°N),seobserva que la temperatura ambiente (Tamb)media en los meses de mayo y junio de 2010 a2015 osciló entre 29,3 y 33,1°C. Esta franja seencuentra en el rango que, según la literatura,favorece ciclos de vida cortos en los triatominos(temperatura superior a 28 °C) (21-24).
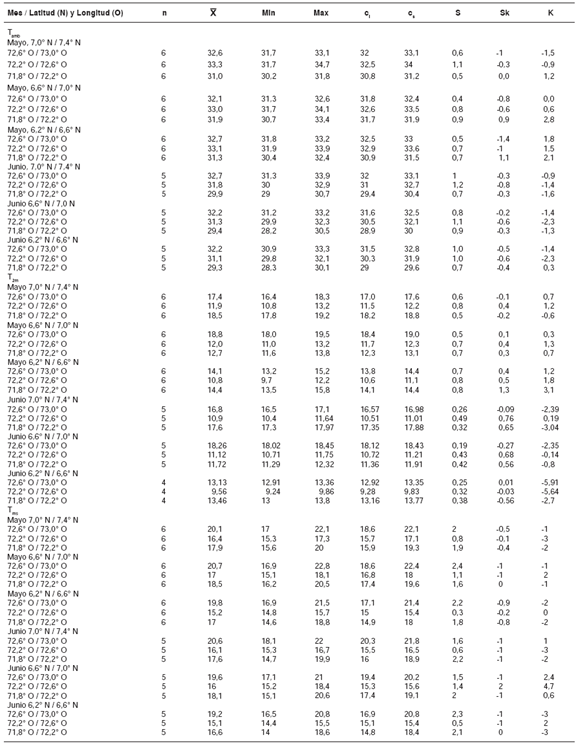
Según el rango de los valores mínimos y máximos,principalmente en los cuartiles inferior y superior,la temperatura ambiente en esta zona y en esos meses es relativamente estable, lo cual satisfacelas necesidades de los triatominos. En general, elmínimo observado fue de 28,3 °C y el máximo fuede 33,9 °C.
Los resultados estadísticos indicaron que el coeficientedeasimetríadelosvaloresdelatemperaturadelmesdemayodelosaños2010a2015seagruparonpordebajodelamediaobservadaparacadaárea, pero nunca por debajo de 28 °C.
Con respecto al mes de junio, los valores se distribuyeronde forma relativamente simétrica enla mayoría de las coordenadas longitudinales(oriente-occidente). En otras palabras, en junio losvalores fueron algo más dispersos que en mayo,lo cual permitiría concluir que, comparativamente, en junio se presentarían ciclos de vida más cortos en los triatominos.
Por otro lado, al comparar los meses de mayo y junio,los resultados indicaron que en el área comprendidaentre los 7,6°N y 73°O, franja en la que se localizael municipio de Soatá (Boyacá), las condicionesambientales en mayo eran más estables para lasnecesidades del ciclo de vida de los triatominos.
El coeficiente de curtosis indicó que, en la mayoríade las franjas longitudinales, la distribución fue muyapiñada (leptocúrtica) en mayo, y fue muy dispersa (platicúrtica) en junio, lo cual concuerda con lainterpretación del coeficiente de asimetría, dado elacentuado grado de concentración de los valoresde la temperatura en mayo y la condición inversaen junio.
Una dispersión similar de los valores de la Tambse observó en las franjas longitudiales del rangolatitudinal entre los 6,6° y los 7,0°N, y en losrangos longitudinales entre los 72,6° y los 73,0°Oy los 73,0° y los 73,4°O, donde se encuentran lascondiciones más estables para los triatominos. Enestas franjas de latitud y longitud, se localizan losmunicipios de Capitanejo, Macaravita, Tipacoque yMolagavita. En la franja longitudinal entre los 73,0°y los 73,4°O, se localiza el municipio de San Vicentede Chucurí, localidad con índices de infestación por triatominos de entre 5 y 7%.
El comportamiento interanual de la media de la Tamb(figura 2a) entre 2010 y 2015 sugiere que en juniohubo una inversión en los valores correspondientesa la franja longitudinal de los 72,6°O con respectoa la de los 73,0°O, en tanto que, en la franjalongitudinal más hacia el oriente de la cordillera,se registraron menores valores en todos los casos. En cuanto a sus características geográficas, esa franja es la más influenciada por la zona de alta montaña del Parque Nacional Natural El Cocuy.
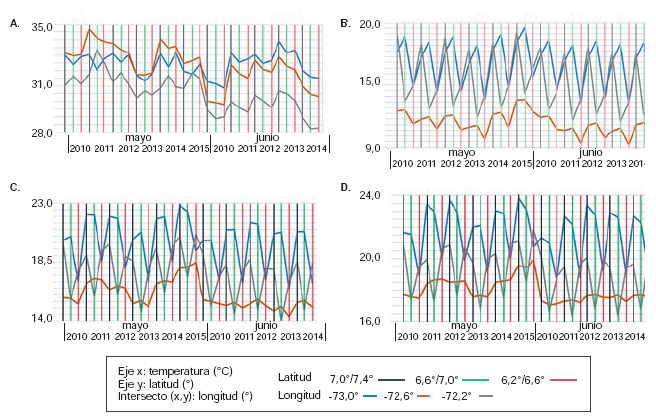
Figura 2
Valores de la Tamb (A), T2m (B), Tms (C) y Tsc (D) en las diferentes franjas latitudinales y longitudinales en los meses de mayo y junio entre 2010 y 2015
En la figura 3 a y b, se observa cómo de mayo ajunio se registró un desplazamiento espacial de losvalores máximos y mínimos de la temperatura mediahacia el oeste de las ecorregiones, lo cual implicauna exposición de los triatominos localizados enla ladera occidental de la Cordillera Oriental y lafrontera norte de la montaña santandereana y eloriente del cañón de los ríos Chicamocha y Suáreza temperaturas algo más bajas; este sector tienelos registros más bajos del vector según la base dedatos utilizada. Una situación inversa se presentócon las poblaciones de triatominos localizados enla porción nororiental del Magdalena Medio, y las localizadas en la región central del cañón, es decir,las áreas en las que los puntos de reporte fueronmás dispersos.
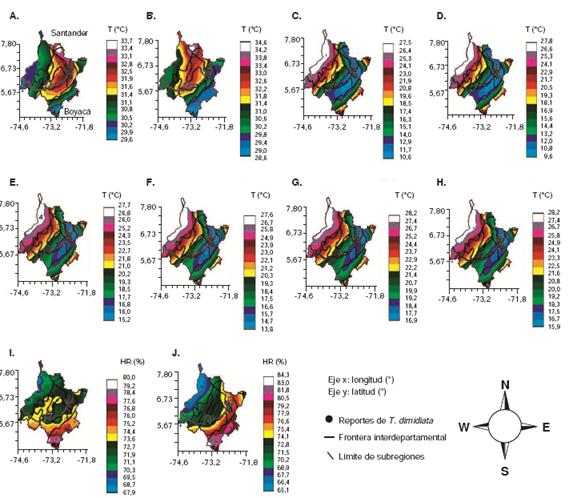
Figura 3
Mapas resultantes de la interpolación por krigeage del promedio de los años 2010 y 2015 para laTamb de mayo (A) y junio (B); la T2m de mayo (C) y junio (D); la Tms de mayo (E) y junio (F); la Tsc de mayo (G) y junio (H). Interpolación por krigeage de la humedad relativa para mayo (I) y junio (J). (1) Ladera occidental de la Cordillera Central; (2) montaña santandereana; (3) cañón de los ríos Chicamocha y Suárez; (4) Magdalena Medio. Los puntos indican los sitios de recolección de T. dimidiata. Distancia de separación en grados decimales en los variogramas isotrópicos. Resolución espacial: 40 km
Temperatura promedio a dos metros de lasuperficie del suelo
Es evidente que la temperatura constituye un parámetrobiofísicodeprimerordenparacomprenderlaecologíadelostriatominos.Hastaahorasehabíalogradoinferirlascondicionesgeneralesdelatemperaturamedia en las áreas donde hay registrode T. dimidiata, es decir, en una escala verticalgrosera, lo que es importante para interpretar el contexto ambiental de los biotopos donde estostriatominos están presentes. Sin embargo, comose puede deducir de los trabajos en campo y en ellaboratorio ya citados, los triatominos responden avariaciones bien localizadas del ambiente.
Por consiguiente, se impone mejorar la resoluciónhorizontal y la vertical de la temperatura, o ambas.En este caso, el modelo escogido cuenta con unaserie de variables que mejoran la resolución verticalde este parámetro, en tanto que el ajuste de la resoluciónhorizontalseplanteaparafuturosestudios. Acontinuaciónsepresentanlosresultadosconformealosvaloresmediosdelatemperaturapromedioados metros del suelo (T2m) en los ecótopos ylocalidades con presencia de T. dimidiata.
En todos los rangos latitudinales y longitudinalesde las áreas de muestreo, fue posible observarvalores medios de temperatura significativamentemás bajos que la media de la temperatura ambientegeneral, tanto en mayo como en junio (cuadro 1b).
La temperatura mínima (9,2 °C) registrada en juniosugiere que esta variable pudo llegar a ser entre19 y 20°C más baja que la mínima registrada parala temperatura ambiente (29,4 °C). Por lo tanto, latemperatura media registrada para la zona másdensamente poblada por vectores (9,6 °C), correspondióa la media más baja registrada entre lasfranjas latitudinales y longitudinales consideradas enel estudio. Estos registros mínimos de la temperaturamedia coincidieron con el mínimo reportado comoel límite de temperatura inferior soportable para lostriatominos de regiones latitudinales más septentrionales,como lo sugieren Galvão, etal.(50).
En la figura 2b se puede observar la variaciónanual de los valores medios de la T2m. En la figurase observa también que: (i) en la franja longitudinalde los 72,2°O y los 72,6°O, siempre se presentó lamenor media de T2m; (ii) en esta misma longitud, lafranja latitudinal más ecuatorial (6,2° y 6,6°N) siemprepresentólamenormediadetemperatura;(iii)enmayosepresentaronlospicos másaltosdeT2m;(iv)enlafranjalatitudinal(6,2°y6,6°N),ladiferenciadetemperaturaentrelafranja(72,2°a72,6°O)ysusvecinas,fue regular y, aproximadamente, 3,5 °Cmenor tanto en mayo como en junio.
En la figura 3, c y d, correspondientes a mayo yjunio, respectivamente, se observa claramente lavariación de la T2m entre las subregiones ecológicasde los departamentos en cuestión. En estosmapas puede verse que la zona de menor T2m media forma lo que se podría llamar una depresión de la temperatura a nivel espacial (áreas en azulde la figura 3, c y d), lo cual coincide en cuanto a suorientación y localización (SO-NO) con la cuencadel río Sogamoso, en un área encañonada quecolinda hacia el noreste con el nudo de Santurbány hacia el este con la Sierra Nevada del Cocuy,cuyas elevaciones superan los 2.500 y 3.000 msnm,en tanto que en su costado oeste puede llegar aelevaciones por encima de los 2.000 msnm.
Temperatura media de la superficie del suelo
Los menores valores medios de la temperaturamedia de la superficie del suelo (Tms) (15,2 °C enmayo y 15,1 °C en junio), se encontraron justamenteen las franjas latitudinal y longitudinal donde sereportó la presencia de T. dimidiata durante laépoca estudiada (latitud: 6,2° y 6,6°N; longitud:72,2° y 72,6°O). Por otro lado, en la franja latitudinaly en la longitudinal (7,0° y 7,4°N; 73,0° y 73,4°O)donde no ha habido reportes de triatominos, o sonmás escasos, se presentaron los mayores valoresmedios de Tms(cuadro 1c).
En la figura 2c se aprecia la fluctuación de la Tms encada franja latitudinal y longitudinal en el periodo de2010 a 2015. Puede constatarse que los menoresvalores de temperatura media se registraron enla franja con más reportes del vector (latitud: 6,2°y 6,6°N; longitud: 72,2° y 72,6°O) tanto en mayocomo en junio.
Para confirmar lo que hasta ahora se ha planteado,puede recurrirse a los datos de la temperatura enel suelo cubierto (Tsc) (figura 2d), los cuales revelanque la Tscmedia fue, por lo menos, 2 °C más alta quela media de la Tms, y en algunas localidades, hasta6 °C más alta,aun cuando siguió prácticamenteel mismo patrón de las fluctuaciones encontradasen el lapso estudiado y en cada franja latitudinal y longitudinal.
La interpolación de los valores según las subregionesecológicasdelosdosdepartamentospermiteverquelavariacióndelaTms(figura2c,d)siguióelmismopatrónespacialdelaTsc(figura2e,f),aunqueconuna media 2 °C más alta, aproximadamente.Este mismo patrón de distribución espacial seobservó en los valores de la T2m, pero a temperaturasmás bajas, lo cual sugiere una variación resultantede estas tres variables a nivel local y regional.
Densidad de la cobertura vegetal y pluviosidad
Se observó que, comparativamente, la densidadpromedio de la cobertura vegetal en las áreas demuestreo de Santander fue mayor que en Boyacá (cuadro 2). Como se mencionó en la sección demateriales y métodos, según la base de datosutilizada no hubo recoleccion del vector domiciliadoen Santander, pues todos los ejemplares recolectadoscorrespondíanalvectorselvático,mientrasqueenlosmunicipiosboyacencessedioelcasoinverso,tanto en los biotopos simplificados ymarcadamente antrópicos (figura 4a) como enaquellos con una cobertura vegetal densa (figura4b), correspondiente a una matriz de bosquesfragmentados, como puede inferirse de los valoresdel EVI2 obtenidos punto a punto (figura 4b).
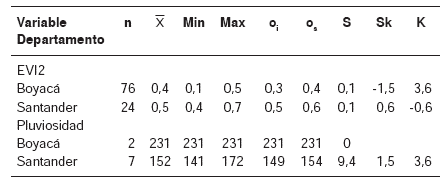
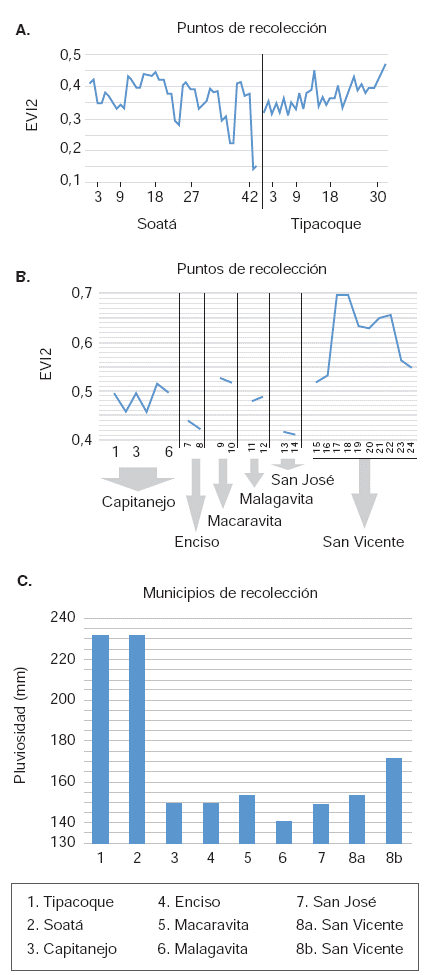
Figura 4
Valores del EVI2 en los puntos de recolección de T. dimidiata en municipios de Boyacá (A) y Santander (B) en mayo y junio de 2007. Valores de (A) y (B) conforme al promedio mensual de imágenes MODIS, producto MOD13, colección 005, resolución espacial de 250 m; valores de (C) conforme al acumulado mensual para ambos departamentos en sensor TRMM, resolución espacial de 5 km
En condiciones normales, la alta pluviosidad en eltrópico se asocia con una mayor densidad de lacobertura vegetal. En este caso, los valores de laprecipitación de los municipios boyacences fueronsignificativamente más altos que los observados en los municipios santandereanos (figura 4c), porlo que cabía esperar valores del EVI2 más altos enlos lugares de muestreo de Boyacá. Se evidencióque los valores del EVI2 en los sitios de muestreoen este departamento correspondían a áreas cuyacobertura vegetal se ha visto sometida a la intervencióndel hombre, transformándose en formasmás simplificadas y menos densas en comparacióncon los lugares de muestreo en Santander,aspecto que se puede confirmar visualmente enlas imágenes satelitales correspondientes.
Análisis armónico de la densidad de lacoberturavegetal y la pluviosidad
Otra situación que se detectó y que vale la penaexplorar, fue el comportamiento de las variablesfísicas y ecológicas a lo largo del año, lo cual esuno de los factores determinantes en la fluctuaciónde las poblaciones de T. dimidiata y que, en elpresente estudio, se expresaron en los armónicosde la densidad, de la precipitación mensual y lacobertura vegetal (EVI2).
Los datos de la pluviosidad y del EVI2 en los diferentes sectores exhibieron un patrón muy similar, con una máxima variación en los periodos de 12 y 24 meses, siendo más predominante este último.
En el cuadro 3, a y b, se presentan la amplitudy las fases de la pluviosidad y de la coberturavegetal, respectivamente. A pesar de que los datosdel muestreo de las distintas localidades fuerondiferentes entre sí, se encontró que los patronesde variación de los datos de pluviosidad fueron losmismos, es decir, en todos los lugares la variaciónde las lluvias siguió los mismos patrones para losperiodos bienal y anual, conforme se observa en lafigura 5, a y b.

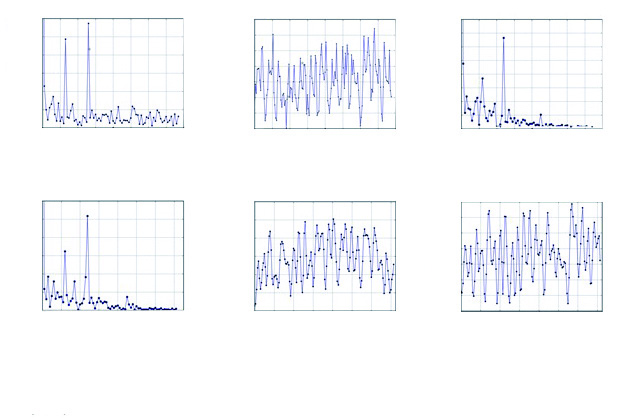
Figura 5
(A) Espectro de amplitudes de la transformada de Fourier en función del tiempo y la pluviosidad. (B) Espectro de amplitudes de la transformada de Fourier en función del tiempo y de los datos de pluviosidad en Capitanejo. (C) Espectro de amplitudes de la transformada de Fourier en función del tiempo y de los datos de pluviosidad en Enciso. (D) Espectro de amplitudes de la transformada de Fourier en función del tiempo y de los datos de pluviosidad en Tipacoque. (E) Espectro de amplitudes de la transformada de Fourier en función del tiempo y de los datos del EVI2 en Enciso. (F) Espectro de amplitudes de la transformada de Fourier en función del tiempo y de los datos del EVI2 en Tipacoque
De los datos del cuadro 3 se desprende que enel período anual se presentó una mayor variaciónalrededor de noviembre y, en el bienal, la máxima sedio en el mes de agosto del segundo año (2002).
Algo similar ocurrió con los índices de coberturavegetal en cada localidad, que si bien no obedecieronexactamente al mismo patrón, registraronla misma tendencia, aunque con diferentes intensidadesde variación en los periodos anualesy bienales, lo cual permite a las poblacionesde T. dimidiata desarrollarse tanto en biotopossimplificados y acentuadamente antrópicos (figura4a) como en biotopos con cobertura vegetal densa(figura 4b). A manera de ejemplo, se presentan doscasos: la localidad de Enciso en Santander y la deTipacoque en Boyacá (figura 5, c, d, e y f).
En el cuadro 3b se observa la fase de los dosperiodos. En Tipacoque, el máximo anual se diohacia el mes de octubre en el periodo bienal, y sumáximo se registró entre noviembre y diciembre delprimer año (2001). En Enciso, se observó un pico amediados de noviembre en su periodo anual, y enel periodo bienal el máximo ocurrió alrededor delmes de mayo del segundo año (2002).
Humedad relativa media
La humedad relativa fue alta en todas las latitudesy longitudes geográficas (cuadro 4) y en losdos meses, inclusive al considerar los mínimosobservados. Los valores, correspondientes alperíodo de 2010 a 2015, fluctuaron entre 66,8 y85,1 %, lo cual indica una alta pluviosidad en lazona, por lo que se asumió que las condicionesclimáticas no difirieron mucho de las encontradasen el 2007, año en que se efectuaron las actividadesdemuestreo.
En la figura 3, i y j, se presenta el comportamiento espacial de la humedad relativa media y puedeobservarse cierta tendencia al incremento en lafranja longitudinal correspondiente a los 72,2° ylos 72,6°O, es decir, aquella donde la temperatura media del ambiente se mostró ligeramente menor, lo cual fue más evidente en los valores correspondientesa junio.
La concentración de T. dimidiata en refugios mássecos podría resultar en la infestación del domicilioa partir de los remanentes de bosques cercanosen áreas que, como la zona de estudio, han estadosometidos a cambios antrópicos. Esto, a su vez,sugiere que puede haber una mayor incidencia enla ingestión de sangre y, consecuentemente, unaumento en el tamaño de las poblaciones. En lasfiguras también se observa un aumento espacialdel porcentaje de humedad relativa entre mayoy junio,sobre todo en las áreas correspondientes a la ladera occidental de la Cordillera Oriental ya la frontera norte de la montaña santandereanay el oriente del cañón de los ríos Chicamocha ySuárez, que, como ya se dijo, es el sector conmenor dispersión de T. dimidiata. Asimismo, seregistró una situación inversa para las poblacioneslocalizadas en la porción nororiental del MagdalenaMedio y en la región central del cañón, es decir, las áreas en las que los puntos de reporte de T.dimidiata aparecen más dispersos. Sin embargo,no se sabe qué tan significativas pueden ser estasdiferencias, ya que incluso los valores mínimos obtenidos con los modelos utilizados clasificarían como de alta humedad relativa.
Discusión
Se observó que la Tamben las áreas de muestreoosciló entre 28,3 y 33,9 °C. Según el sistemade Köppen (27), el clima de la región puedeconsiderarse como macrotérmico. Las variablesanalizadas, más otras de tipo biótico, como la disponibilidadde hábitat y de fuentes de alimento,reflejaron condiciones estables que aumentan laprobabilidad de una amplia dispersión de la especiey favorecen su mantenimiento y crecimiento.
Diversos autores (51-57) han señalado que es deesperar que los ciclos de vida sean más cortos a temperaturas constantes que oscilen entre 27y 28 °C que aquellos de colonias mantenidasa temperaturas variables. En este sentido, losvalores observados se mantuvieron en ese rangoen mayo y junio, por lo menos durante los últimoscinco años del periodo de estudio, por lo cual cabeesperar que tales condiciones se hayan mantenidopara las poblaciones de triatominos.
Estos resultados a nivel macroclimático aparentementeconcuerdan con los obtenidos en ellaboratorio por diversos autores para diferentesespecies de triatominos. En esas condiciones, laduración media del ciclo de vida a 25 °C siempre hasido mayor que a 30 °C (55,57-60). Con respecto aT. dimidiata, Zeledón, et al. (58), reportaron que loshuevos incubados a temperaturas de 22 a 24 °Cnecesitaron un lapso más largo para eclosionar (29días) que los incubados a 26 °C (23 días).
No obstante, los resultados obtenidos para las variablesde temperatura (T2m, Tms y Tsc) indican que,según lo han sugerido varios autores, en realidadlas condiciones distaron de ser las más favorables para los triatominos (18-28). Por ejemplo, en el áreacon más registros de T. dimidiata, la Tmsosciló entre14,57 y 18,78 °C en mayo, y entre 14,37 y 15,55 °Cen junio, es decir, una diferencia en la Tamb de hasta13,93 °C en los valores mínimos y de 15,2 °C en los máximos. Es decir que, según los valores obtenidosen las variables T2m, Tms y Tsc, debe esperarse quelos ciclos de vida de las poblaciones de T. dimidataestudiadas fueran largos, a pesar de que los valoresde la temperatura ambiente sugieran lo contrario.De verificarse esto, debe aceptarse que la T2m, laTms y la Tsc serían variables más apropiadas que laTamb para conocer las condiciones de temperatura insitu que soportan los triatominos.
Dichos resultados explicarían por qué varios autoreshan afirmado que los límites de temperaturasuperior e inferior soportables para la supervivenciade colonias de triatominos en condiciones naturalesesrelativamenteamplio.Porejemplo,paraT.infestans,estoslímitesseencuentranentrelos10ylos 37 °C (53) y, para las especies argentinas, el mínimo necesario para su alimentación, su dispersión y su reproducción es de 20 °C (22,59).Así las cosas, es probable que estos valores noreflejen lo que ocurre con las poblaciones detriatominos en condiciones naturales.
Por ejemplo, Lorenzo, et al. (60), hallaron que, encondiciones de laboratorio, T. infestans prefiererefugios a una temperatura de 34 °C. Sin embargo,estos mismos autores sugieren que, bajo condicionesclimáticasnaturales,algunostiposdedomicilioseríanrefugiosfavorablesparaestaespeciedebidoaqueofrecenlascondicionesclimáticasadecuadasyestables, pues sirven de amortiguador frente alas fluctuaciones entre las temperaturas mínimasy máximas.
Curto de Casas, et al. (22), sugieren que elfactor climático importante es el número de díascon temperaturas superiores a 20 °C y no tantola temperatura más baja, como se suponía. Sinembargo, de incluirse las variables T2m, Tms y Tsc,se esperaría que fuera más importante el númerode días con temperatura estable, aunque nonecesariamente superior a 20 °C. Debe resaltarseque la mayoría de los autores hacen referencia acondiciones de laboratorio o temperatura ambiente(25,26), por lo que en este estudio, las observacionesdelaT2m,laTmsylaTscsepostulancomoinéditas,por lo menos en lo que se refiere a T.dimidiata. Dado que son variables de resoluciónprincipalmente vertical, y teniendo en cuenta queen los triatominos no predomina el hábito del vuelo,se sugiere que estas son más adecuadas paraentender el entorno ecológico de esta especie.
Por definición, la Tms depende de la cantidad deradiación neta, lo cual resulta del equilibrio energéticoentreondascortasyondaslargas,ydepende de factores extrínsecos, como la radiación globaldisponible, el albedo y el balance resultante de laradiación infrarroja. A su vez, esta última dependede la temperatura ambiente y de la capacidad deemisión de la atmósfera y la tierra (61).
Como se mencionó en la sección de resultados,los valores más bajos de Tms se presentaron enlas áreas donde más se reportó la presencia deT. dimidiata (con temperaturas entre los 14,6 y los18,8 °C en mayo, y entre los 14,4 y los 15,6 °Cen junio), lo cual respalda las deducciones sobrelas condiciones ecológicas y ambientales localesque, ligadas a las macroclimáticas, incentivanla domiciliación. Sin embargo, por sí solo, estono necesariamente significa que con valoresmáximos de temperatura en la superficie del suelose deba descartar la presencia de triatominosen esas zonas. Hay que recordar que en lospuntos de recolección en esta franja latitudinaly longitudinal se encontraron ejemplares de T.dimidiata domiciliados, por lo que, aparentemente,la mayor concentración del vector en las áreascuya superficie del suelo es más fría coincide conla inferencia hecha con la variable anterior (T2m).En otras palabras, los resultados sugieren queuna temperatura más baja a nivel de la superficiedel suelo, al igual que la T2m, incentiva al vector abuscar refugios (que ya de por sí han sido sustituidospor los artificiales, con temperaturas máscálidas y estables), y una fuente de alimento demenor costo energético.
El significado ecológico de la T2m para los triatominosesalgoincierto.Enregionesboreales,algunosautoresequiparanlaT2mcomolatemperaturaeneldosel(51),peroenlasregionestropicalesnohayinformaciónexactaenestesentido. LaestructuraylafuncióndelaT2mdependendeloscontrastesenlaspropiedadesdelasuperficiedelsuelo(porejemplo,entretierraymar),delrelieve,laelevaciónaltitudinal(62)ylaposiciónlatitudinal.Conrespectoaestavariable,yconbaseenunmuestreorealizadoendiversoslugaresdeSuramérica,Pereira,etal.(63),encontraronquelasustitucióndeáreasconvegetaciónarbóreayotrostiposdeusodelsuelopuedeacarrearunaumento dehasta2°Cen laT2m.
Como ya se mencionó, normalmente se esperaque el vector se localice más a nivel del suelo,entre cuevas y rocas, pero también en oquedadesy nidos de vertebrados, los cuales pueden perfectamenteencontrarse a esta altura del suelo,aunque dichos refugios les permiten evitar laexposición al ambiente externo. Esto sugeriría, por lo menos, tres posibilidades: primero, la T2men esta región puede estar restringiendo el nichode estas poblaciones a ecótopos más a nivelde suelo, donde la temperatura es más alta;segundo, el vector bien puede ocupar ecótoposya mencionados a esta altura del suelo, donde lascondiciones microclimáticas son más favorables,es decir, la temperatura es mayor a la externa, ytercero, una combinación de las dos posibilidadesanteriores, lo que resulta más probable.
Desde el punto de vista epidemiológico, cualquierade estas situaciones puede aparecer cuandose le suman los problemas de deforestación enla región. La transformación del espacio no soloincluye la desaparición de la vegetación nativa,también implica la aparición de nuevos ecótopos que pueden cumplir igual función, o mejor, que los originales, pues las condicionesmacroclimáticasextremas a este nivel del suelo ya no tienenun papel importante para la regulación de laspoblaciones locales de T. dimidiata, pues este locumplen las viviendas humanas y la infraestructuracreada para las actividades económicas o desustento, como gallineros, marraneras, etc., quese pueden considerar como ecótopos artificiales,equivalentes desde el punto de vista funcional alos refugios naturales.
Curto de Casas, et al. (22), también sugieren que,desde el punto de vista macroclimático, una altahumedad del ambiente incrementa la distribucióngeográfica de especies tropicales y subtropicalespropias de los bosques húmedos o inundables; porotro lado, una baja humedad disminuye la duracióndel ciclo de vida de especies propias de bosquessecos tropicales y subtropicales y de áreas áridas,al tiempo que se incrementa su distribución geográficaysudensidadpoblacional.Losresultadosobservadosenestetrabajoconcuerdanconestasaseveraciones.
En este estudio se observó que la humedad relativaoscilaba entre 66,8 y 85,6 %. Lorenzo, et al. (60),encontraron que, en condiciones de laboratorio, T.infestans prefiere refugios con humedad relativade alrededor de 20 %. Al igual que la temperaturaen las viviendas, algunos materiales usados enla construcción de techos, como la jarilla (Larreasp.), amortiguan las fluctuaciones entre los valoresmínimos y máximos de humedad relativa.
Por otro lado, Xavier, et al. (64), sugieren que,en condiciones de laboratorio, los ejemplares deT. brasiliensis recién alimentados pueden buscaractivamente áreas dentro de sus refugios donde la humedad relativa sea baja, con el objetivo deperder su exceso de agua sin costo metabólico, oadoptar un comportamiento opuesto en situacionesde estrés hídrico.
La humedad relativa observada en este estudio es ladel ambiente en general, por lo que es posible que,a nivel microclimático, en realidad se presenten lascondiciones observadas por los autores citados, yno las generales, tal y como se observó con lasvariables de temperatura.
En este sentido, las observaciones hechas eneste trabajo sugieren que una alta humedadrelativa genera en los ejemplares de T. dimidiatarecién alimentados un comportamiento similar alencontrado en T. brasiliensis, es decir, la búsqueday permanencia en refugios más secos ycon temperaturas más bajas, lo que en la prácticaconvierte a los domicilios humanos en refugiosinmejorables.
Por lo menos desde el punto de vista de losvalores de temperatura y humedad relativa enlas ecorregiones involucradas, se evidenció unarelación inversa entre las dos variables, lo cualpuede ser suficiente para desencadenar procesosbiológicos y de comportamiento en los triatominosque impliquen una variación en el riesgo de infestaciónde las zonas pobladas.
En cuanto a la densidad de la cobertura vegetal,se evidenció que los valores de EVI2 observadosen los sitios de muestreo en Boyacá correspondíana áreas cuya cobertura vegetal se ha visto muyafectada por intervenciones externas, transformándolosenáreasmássimplificadasymenosdensas,encomparación con los lugares de muestreo enSantander. Ello sugiere que la domiciliación de T.dimidiata no ocurre obligadamente cuando aún existe en las proximidades una cobertura vegetalnatural y los ecótopos correspondientes, y esun buen indicio de la capacidad de adaptacióny la resiliencia de T. dimidiata frente a diversascondiciones ecológicas y ambientales, lo cual lepermite desenvolverse en biotopos simplificados ymuy antrópicos (figura 4a), así como en aquellosubicados en lugares con cobertura vegetal densa(figura 4b).
Sin embargo, esto no necesariamente significa quelas áreas con vegetación menos densa favorezcanlas poblaciones de T. dimidiata. Debe considerarseque, en cierta forma, los triatominos selváticospueden estar espacialmente más dispersos, lo queen términos biológicos resulta en una disminución de la probabilidad de cruces consanguíneos. Además,es posible que T. dimidiata mantenga sucondición selvática en áreas que, de hecho, notienen una estación seca bien definida, lo cual yase ha evidenciado en especies como Panstrongylusmegistus y T. sordida en el hemisferio sur (65), yen Rhodnius prolixus en el río Orinoco (66).
La condición selvática favorece la heterozigosis, auncuando, en general, se acepta que los miembros deesta subfamilia son gregarios y en términos de laecología de las poblaciones se habla de su dispersióno emigración cuando las colonias son muydensas y logran afectar e irritar a los huéspedes,aumentando la competencia entre ellas.
En términos biológicos, la subsecuente concentracióndelapoblaciónenlosdomiciliossugiereunaumentodelaprobabilidaddecrucesendogámicos,esdecir,unfavorecimientodelahomozigosis,loquepuedesignificar,alargoplazo,undeterioroenlacapacidaddereaccióndelvectoracondicionesadversas. Desde este punto de vista, habría quepreguntarse si la domiciliación de T. dimidiata en lazona puede deberse realmente a una preferenciade hábitat de la especie, o es la manifestación deuna reducción del hábitat natural disponible parasus poblaciones.
Desde el punto de vista epidemiológico, las consecuenciasson otras. Obviamente, una mayordisponibilidad de vectores en espacios reducidos,como es el caso de los insectos domiciliados,implica una mayor probabilidad de difusión deT. cruzi en la población y, por ende, una mayorprobabilidad de transmisión de la enfermedad a loshumanos que conviven con el vector.
Con base en el análisis de las series temporales,se concluyó que durante el tiempo de muestreo, poco más de diez años, la tendencia bienal tuvo mayor dominancia que la anual en el marco de latendencia general del periodo dominante.
Desde el punto de vista de los patrones anual ybienal de la pluviosidad, si se acepta que una altahumedad del ambiente incrementa la distribucióngeográfica de las especies tropicales y subtropicalesde los bosques húmedos, podría esperarse que ennoviembre de cada año se produzca un aumentoen la distribución de los ejemplares de T. dimidiatano domiciliados, y que en el período bienal esteaumento se presente en el mes de agosto de losaños pares. Estos hallazgos pueden servir parafocalizar los periodos de vigilancia entomológicaen la región.
En relación con la densidad de la cobertura vegetal,su aumento en mayo prodría favorer las poblacionesde vectores en relación con las condicionesbioclimáticas relacionadas, por ejemplo, la temperaturaaniveldelsuelo,auncuandonotantoentérminosde humedad del ambiente.
La enfermedad de Chagas es una zoonosis muycompleja que representa una grave amenaza parala salud pública. En ausencia de una vacuna y deun adecuado tratamiento médico, además de lasubvaloración de la magnitud de la enfermedad(se estima que más de 70 % de los pacientesinfectados no son diagnosticados), los principalesmecanismos para su control y eliminación debenbasarse en medidas preventivas. La estrategia queha demostrado mayor efectividad en la lucha contrala enfermedad de Chagas es el control vectorial, conel cual se pretende reducir el número de insectosantropofílicos mediante el uso específico, controladoyefectivodeinsecticidasenlasviviendas.Sinembargo,estas medidas de interrupción verticalsolo han sido efectivas en zonas donde los insectosvectores se encuentran domiciliados, aunque se hanreportado nuevas colonizaciones recurrentemente.En Colombia cada vez se reportan más triatominosy mamíferos intrusos en las viviendas, y recientementese han presentado brotes de enfermedadaguda relacionada con dicha intrusión. En estesentido, se sugiere que las estrategias de controlvectorial empleadas actualmente sean diseñadasy ejecutadas de manera local y se ajustena las características biológicas de las especiesde triatominos y las condiciones epidemiológicasen cada caso, por lo cual, para las poblacionessecundarias e intrusas de vectores, las alternativasque han mostrado mayor efectividad para elcontrol demandan un conocimiento cabal de estas características.
En el esquema clásico de manejo y control de laenfermedad, el significado ecológico y biológico dela domiciliación no ha recibido la debida atenciónhasta ahora; en este estudio se sugiere que estaes adversa a largo plazo para las poblaciones devectores. Este punto de vista permite un nuevoenfoque del problema epidemiológico, el cual ya seha tenido en cuenta para el control químico, puesse evidenció que, debido a la homogeneidad genética de la especie, han sido pocos los hallazgos de resistencia a insecticidas.
Hasta hoy, los programas de control y manejo dela enfermedad se han enfocado en medidas contingentesbasadas,sobretodo,enlaeliminacióndel vector domiciliado; sin embargo, estos programasno han podido impedir nuevas infestaciones delvector selvático.
En el marco del enfoque ecoepidemiológico delproblema, se sugiere adoptar medidas contingentesy preventivas contra el vector no domiciliado, puessu presencia entraña nuevas posibilidades de transmisiónquedebenabordarsedemaneradiferente,sobretodo en el entendido de que los ciclos derociado con insecticidas piretroides no logran evitarlas nuevas infestaciones. Este, que es el métodoactual, aparentemente preventivo, en realidad esevidentemente contingente y, por ende, puedecuestionarse su efectividad para disminuir el riesgode nuevas infestaciones de vectores selváticos.
En todo caso, desde una perspectiva ecológicaresulta difícil garantizar la erradicación real y permanentedelvectorsinoseavanzaenelconocimientodelvector no domiciliado y, por supuesto, en eldiseño de estrategias para su control.
Por último, hay suficiente información que permitiríael modelado y la proyección de la distribución delos triatominos, y de T. dimidiata en particular. Losmodelos propuestos por diversas universidades,disponibles en los portales del IPCC, permitirían,una vez ajustados a las políticas de estado, obtenerun panorama fiable del futuro de la situación delos vectores y del contexto epidemiológico de laenfermedad de Chagas en Colombia.
Agradecimientos
A la Red Chagas Colombia y a Skandha EIT
Referencias
1. Dumonteil E, Gourbie?re S, Barrera-Pérez M, RodríguezFélix, Ruiz-Piña H, Baños-López O, et al. Geographic distribution of Triatoma dimidiata and transmission dynamics of Trypanosoma cruzi in the Yucatán peninsula of México. Am J Trop Med Hyg. 2002;67:176-83. https://doi.org/10.4269/ajtmh.2002.67.176
2. Guhl F, Aguilera G, Pinto N, Vergara D. Actualización de la distribución geográfica y ecoepidemiología de la fauna de triatominos (Reduviidae:Triatominae) en Colombia. Biomédica. 2007;27:143-62. https://doi.org/10.7705/biomedica.v27i1.258
3. Calderón CI, Dorn PL, Melgar S, Chávez JJ, Rodas A, Rosales R, et al. A preliminary assessment of genetic differentiation of Triatoma dimidiata (Hemiptera: Reduviidae) in Guatemala by random amplification of polymorphic DNApolymerase chain reaction. J Med Entomol. 2004;41:882-7. https://doi.org/10.1603/0022-2585-41.5.882
4. Schofield CJ. Evolución y control del Triatoma dimidiata En: Organización Panamericana de la Salud, editor. Taller para el establecimiento de pautas técnicas en el control de Triatoma dimidiata. (Documento OPS/HCP/HCT/214/02). San Salvador: Organización Panamerica de la Salud; 2002.p. 12-8.
5. Dorn PL, Monroy C, Curtis A. Triatoma dimidiata (Latreille, 1811): A review of its diversity across its geographic range and the relationship among populations. Infect Gen Evol. 2007;7:343-52. https://doi.org/10.1016/j.meegid.2006.10.001
6. Blandón-Naranjo M, Zuriaga MA, Azofeifa G, Zeledón R, Bargues MD. Molecular evidence of intraspecific variability in different habitat-related populations of Triatoma dimidiata (Hemiptera: Reduviidae) from Costa Rica. Parasitol Res. 2010;106:895-905. https://doi.org/10.1007/s00436-010-1762-9
7. Bargues MD, Klisiowicz DR, González-Candelas F, Ramsey JM, Monroy C, Ponce C, et al. Phylogeography and genetic variation of Triatoma dimidiata, the main Chagas disease vector in Central America, and its position within the genus Triatoma. PLoS Negl Trop Dis. 2008;7;2:e233. https://doi.org/10.1371/journal.pntd.0000233
8. Monroy MC, Bustamante DM, Rodas AG, Enríquez ME, Rosales RG. Habitats, dispersion and invasion of sylvatic Triatoma dimidiata (Hemiptera: Reduviidae: Triatominae) in Petén, Guatemala. J Med Entomol. 2003;40:800-6.
9. Dorn PL, Calderón C, Melgar S, Moguel B, Solórzano E, Dumonteil E, et al. Two distinct Triatoma dimidiata (Latreille, 1811) taxa are found in sympatry in Guatemala and México. PLoS Negl Trop Dis. 2009;3:e393. https://doi.org/10.1371/journal.pntd.0000393
10. Gómez-Palacio A, Triana O, JaramilloON, Dotson EM, Marcet PL. Eco-geographical differentiation among Colombian populations of the Chagas disease vector Triatoma dimidiata (Hemiptera: Reduviidae). Infect Genet Evol. 2013;20:352-61. https://doi.org/10.1016/j.meegid.2013.09.003
11. Gaunt M, Miles M. The ecotopes and evolution of triatomine bugs (Triatominae) and their associated trypanosomes. Mem Inst Oswaldo Cruz. 2000;95:557-65. https://doi.org/10.1590/S0074-02762000000400019
12. Parra-Henao G. Triatoma dimidiata en Colombia. Distribución, ecología, análisis de factores de riesgo y generación de mapas predictivos de riesgo (tesis). Medellín: Universidad de Antioquia; 2012.
13. Miles M, De Souza A, Povoa M. Chagas' disease in the Amazon basin III. Ecotopes of ten triatomine bug species (Hemiptera: Reduviidae) from the vicinity of Belem, Para State, Brazil. J Med Entomol. 1981;18:266-78.
14. Noireau F, Carbajal-deLaFuente AL, Lopes CM, Diotaiuti L. Some considerations about the ecology of Triatominae. An Acad Bras Cienc. 2005;77:431-6. https://doi.org/10.1590/S0001-37652005000300006
15. Parra-Henao G, Angulo V, Jaramillo N, Restrepo IM. Triatominos (Hemiptera: Reduviidae) de la Sierra Nevada de Santa Marta, Colombia: aspectos epidemiológicos, entomológicos y de distribución. CES Med. 2009;23:17-26.
16. Quiroz-Gómez O, Jaramillo N, Angulo VM, Parra-Henao G. Triatoma dimidiata en Colombia: distribucion, ecología e importancia epidemiológica. Biomédica. 2017;37:274-85. https://doi.org/10.7705/biomedica.v37i2.2893
17. Zeledón R, Montenegro VM, Zeledón O. Evidence of colonization of man-made ecotopes by Triatoma dimidiata (Latreille, 1811) in Costa Rica. Mem Inst Oswaldo Cruz. 2001;96:65960. https://doi.org/10.1590/S0074-02762001000500012
18. Carcavallo RU. Climatic factors related to Chagas disease transmission. Mem Inst Oswaldo Cruz. 1999;94:367-9. https://doi.org/10.1590/S0074-02761999000700071
19. Dumonteil E, Gourbie?re S. Predicción de la abundancia y tasa de infección de Triatoma dimidiata: un mapa de riesgo de transmisión natural de la enfermedad de Chagas en la Península de Yucatán, México. Rev Biomed. 2004;15:221-31.
20. Hack W. Estudios sobre biología del Triatoma infestans (Klug, 1834) (Hemiptera, Reduviidae). An Inst Med Regional. 1955;4:125-47.
21. Carcavallo RU. Aspects of the epidemiology of Chagas disease in Venezuela and Argentina. En: New approaches in American Trypanosomiasis Research. Scientific publication 318. Washington, D.C.: OPS; 1976. p. 318-20.
22. Curto de Casas S, Carcavallo R, Mena Segura C, GalíndezGirón I, Burgos J. Bioclimatic factors of Triatominae distribution. Useful techniques for studies on climatic change. Entomol Vect. 1994;1:51-68.
23. Forattini O, Santos J, Ferreira O. Aspectos ecológicos da tripanossomíase americana. XVI. Dispersão e ciclos anuais de colônias de Triatoma sordida e de Panstrongylus megistus espontaneamente desenvolvidas em ecótopos artificiais. Rev Saúde Pública. 1979;13:299-313. https://doi.org/10.1590/S0034-8910197900040000
24. Forattini OP, Ferreira OA, da Rocha e Silva EO, Rabello EX. Aspectos ecológicos da tripanossomíase americana: XIV - Persistência e potencial de domiciliação de populações triatomínicas silvestres em região de intensa atividade agropecuária. Rev Saúde Pública. 1979;13:123-46.
25. IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press; 2013.
26. Mendonça F, Danni-Oliveira IM. Climatologia: noções básicas e climas do Brasil. São Paulo: Oficina de Textos;2007.
27. Kottek M, Grieser J, Beck C, Rudolf B, Rubel F. World map of the Köppen-Geiger climate classification updated. Meteorologische Zeitschrift. 2006;15:259-63. https://doi.org/10.1127/0941-2948/2006/0130
28. Parra-Henao G. Sistemas de información geográfica y sensores remotos. Aplicaciones en enfermedades transmitidas por vectores. CES Med. 2010;24:75-90.
29. Hernández J, Núñez I, Bacigalupo A, Cattan PE. Modeling the spatial distribution of Chagas disease vectors using environmental variables and people's knowledge. Int J Health Geogr. 2013;12:29. https://doi.org/10.1186/1476-072X-12-29
30. Pfeiffer D, Hugh-Jones M. Geographical information systems as a tool in epidemiological assessment and wildlife disease management. Rev Scient Tech. 2002;21:91-102.
31. Young SG, Tullis JA, Cothren J. A remote sensing and GIS-assisted landscape epidemiology approach to West Nile virus. Applied Geography. 2013;45:241-9. https://doi.org/10.1016/j.apgeog.2013.09.022
32. Rytkönen MJ. Not all maps are equal: GIS and spatial analysis in epidemiology. Int J Circumpolar Health. 2004;63:9-24.
33. Graham AJ, Atkinson PM, Danson FM. Spatial analysis for epidemiology. Acta Trop. 2004;91:219-25. https://doi.org/10.1016/j.actatropica.2004.05.001
34. National Research Council (US) Committee on Environmental Epidemiology. Environmental Epidemiology: Volume 1: Public Health and Hazardous Wastes Washington, D.C.: National Academies Press (US); 1991. p. 1-20.
35. Lai P-C, So F-M, Chan K-W. Spatial epidemiological approaches in disease mapping and analysis. Int J Health Geogr. 2009;8:14. https://doi.org/10.1186/1476-072X-8-14
36. Pfeiffer D, Robinson TP, Stevenson M, Stevens KB, Rogers DJ, Clements AC. Spatial analysis in epidemiology. New York (NY): Oxford University Press; 2008. p. 142. https://doi.org/10.1186/1756-3305-2-23
37. Mateo RG, Felicísimo ÁM, Muñoz J. Modelos de distribución de especies: una revisión sintética. Rev Chil Hist Nat. 2011;84:217-40. https://doi.org/10.4067/S0716-078X2011000200008
38. Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, et al. Novel methods improve prediction of species' distributions from occurrence data. Ecography. 2006;29:129-51. https://doi.org/10.1111/j.2006.0906-7590.04596.x
39. Elliott P, Wartenberg D. Spatial epidemiology: Current approaches and future challenges. Environ Health Perspect. 2004;112:998-1006. https://doi.org/10.1289/ehp.6735
40. Emmanuel NN, Loha N, Okolo MO, Ikenna OK. Landscape epidemiology: An emerging perspective in the mapping and modelling of disease and disease risk factors. Asian Pac J Trop Dis. 2011;1:247-50. https://doi.org/10.1016/S2222-1808(11)60041-8
41. Phillips SJ, Anderson RP, Schapire RE. Maximum entropy modeling of species geographic distributions. Ecol Model. 2006;190:231-59. https://doi.org/10.1016/j.ecolmodel.2005.03.026
42. Bloomfield P. Fourier analysis of time series: An introduction. New York: John Wiley & Sons, Inc.; 2004.
43. De Castro Victoria D, Andrade RG, da Paz AR. Série temporal de imagens EVI/MODIS para discriminação de formações vegetais do Pantanal. Geografia. 2011;34:721-9.
44. Jakubauskas ME, Legates DR, Kastens JH. Harmonic analysis of time-series AVHRR NDVI data. Photogrammetric Engineering and Remote Sensing. 2001;67:461-70.
45. Grupo Intergubernamental de Expertos sobre Cambio Climático. Informe especial del IPCC, escenarios de emisiones. Fecha de consulta: 15 de febrero de 2016. Disponible en: https://www.ipcc.ch/pdf/special-reports/spm/sres-sp.pdf.
46. Gordon C, Cooper C, Senior CA, Banks H, Gregory JM, Johns TC, et al. The simulation of SST, sea ice extents and ocean heat transports in a version of the Hadley Centre coupled model without flux adjustments. Climate Dynamics. 2000;16:147-68. https://doi.org/10.1007/s003820050010
47. Jiang Z, Huete AR, Didan K, Miura T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens Environ. 2008;112:3833-45. https://doi.org/10.1016/j.rse.2008.06.006
48. Jesus PH, Rocha JC. Análise comparativa dos índices de vegetação NDVI e EVI2 na identificação de culturas de cana-de-açúcar em imagens Landsat-5. An Cong de Inic Cien. 2011;7:1-5.
49. Hsu HP, Ranjan R. Signals and systems. New York: McGraw-Hill; 2014.
50. GalvãoC, da Silva RochaD, CunhaV, PresgraveOA, JurbergJ, CarcavalloR. Influence of the temperature in the life cycle of Triatoma melanosoma Martínez, Olmedo & Carcavallo, 1987 (Hemiptera, Reduviidae). Mem Inst Oswaldo Cruz. 1999;94:851-4. https://doi.org/10.1590/S0074-02761999000600027
51. De Roy W, De Vries H. High resolution limited area model (HIRLAM). Harmonie verification and evaluation. Hirlam Technical Report. 2017;70:1-84.
52. Garcia da SilvaI, Garcia da SilvaHH. The influence of temperature on the biology of triatominae: IX. Rhodnius nasutus Stal, 1859 (Hemiptera, Reduviidae). Mem Inst Oswaldo Cruz. 1989;84:377-82. https://doi.org/10.1590/S0074-02761989000300013
53. Carcavallo RU, Martínez A. Life cycles of some species of Triatoma (Hemiptera: Reduviidae). Can Entomol. 1972;104:699-704.
54. Rocha DS, Jurberga J, Carcavallo RU, Presgraveb OA, Cunhaa V, Galvãoa C. Influência da temperatura e umidade no desenvolvimento ninfal de Rhodnius robustus. Rev Saúde Pública. 2001;35:400-6. http://dx.doi.org/10.1590/S0034-89102001000400011
55. Garcia da Silva I, Fernandes FF, Garcia da Silva HH. Influência da temperatura na biologia de triatomíneos. XX. Triatoma maculata (Erichson, 1848) (Hemiptera, Reduviidae). Rev Pat Trop. 1995;24:49-54.
56. Garcia da Silva I, Garcia da Silva HH. Influência da temperatura na biologia de Triatomíneos. III. Panstrongylus megistus (Burmeister, 1835) (Hemiptera, Reduviidae). Rev Bras Entomol. 1988;37:489-96.
57. Garcia da Silva I, Garcia da Silva HH. The influence of temperature on the biology of triatomine. IX. Rhodnius nasutus Stal, 1859 (Hemiptera, Reduviidae). Mem Inst Oswaldo Cruz. 1989;84:377-82. https://doi.org/10.1590/S0074-02761989000300013
58. Zeledón R, Guardia VM, Zúñiga A, Swartzwelder JC. Biology and ethology of Triatoma dimidiata (Latreille, 1811): I. Life cycle, amount of blood ingested, resistance to starvation, and size of adults. J Med Entomol. 1970;7:313-9.
59. Curto de Casas S, Carcavallo R. Límites del triatomismo en la Argentina. I: Patagonia. Chagas. 1984;1:35-40.
60. Lorenzo MG, Lazzari CR. Temperature and relative humidity affect the selection of shelters by Triatoma infestans, vector of Chagas disease. Acta Trop. 1999;72:241-9. https://doi.org/10.1016/S0001-706X(98)00094-1
61. Li Z-L, Tang B-H, Wu H, Ren H, Yan G, Wan Z, et al. Satellite-derived land surface temperature: Current status and perspectives. Remote Sens Environ. 2013;131:14-37. https://doi.org/10.1016/j.rse.2012.12.008
62. Sattler K, Huang XY. Structure function characteristics for 2 meter temperature and relative humidity in different horizontal resolutions. Tellus. 2002;54:14-33. https://doi.org/10.1034/j.1600-0870.2002.00281.x
63. Pereira G, Silva MES, Moraes EC. O uso de produtos orbitais para a parametrização de processos físicos em modelos numéricos regionais e suas alterações climatológicas. Rev Bras Clim. 2012;11:7-24.
64. Xavier AAP, Lorenzo MG, Lazzari C, Diotaiuti L, Guarneri AA. Relative humidity and water loss in Triatoma brasiliensis. Physiolog Entomol. 2005;30:338-42. https://doi.org/10.1111/j.1365-3032.2005.00467.x
65. Forattini OP, Ferreira OA, da Rocha e Silva EO, Rabello EX. Aspectos ecológicos da Tripanossomíase americana: XII - Variação regional da tendência de Panstrongylus megistus à domiciliação. Rev Saúde Pública. 1978;12:20933. https://doi.org/10.1590/S0034-89101978000200013.
66. Gómez-Núñez J. Notas sobre la ecología del Rhodnius prolixus. Bol Malar San Amb. 1963;3:330-5