Soil organic carbon and total nitrogen in a Leucaena leucocephala silvopastoral system in the Chaco region, Argentina
Soil organic carbon and total nitrogen in a Leucaena leucocephala silvopastoral system in the Chaco region, Argentina
RIA. Revista de Investigaciones Agropecuarias, vol. 51, núm. 3, pp. 162-170, 2025
Instituto Nacional de Tecnología Agropecuaria

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Abstract: The introduction of leucaena (Leucaena leucocephala) in silvopastoral systems increases animal production and improves soil fertility. However, there is limited information on the dynamics of carbon and nitrogen in these systems over the long-term periods and soil profile. This study compared the concentrations and vertical distribution of organic carbon (OC) and total nitrogen (TN), and its fractions (particulate and associated forms) in the soil profile (0-100 cm), with and without leucaena, 10 years after its establishment in a tropical pasture (Urochloa brizantha-Chloris gayana). Leucaena increased OC by 73% in the deeper horizon (50-100 cm), particularly in its stable form. It also increased TN by 12% in the surface layer (0-20 cm) and by 21% in the deeper horizon. These increases in OC and TN with leucaena can be associated to root biomass allocation deeper into the soil profile, increased N inputs from leaf deposition, animal manure and urine, and nodule N-fixation. Despite the rise in TN in the surface soil, OC decreased in this layer, which is linked to the low grass cover, a result of high grazing pressure aimed at controlling leucaena hedgerow height. Under proper grazing management, leucaena has the potential to improve soil fertility and nitrogen availability for companion grasses, and may also serve as a greenhouse gas mitigation strategy.
Keywords: legume tree introduction, soils organic carbon, total nitrogen.
Resumen: La introducción de leucaena (Leucaena leucocephala) en sistemas silvopastoriles aumenta la producción animal y mejora la fertilidad del suelo. Sin embargo, hay poca información sobre la dinámica del carbono y nitrógeno en estos sistemas a largo plazo. Este estudio comparó las concentraciones y distribución del carbono orgánico (CO) y nitrógeno total (NT), y sus fracciones (formas particuladas y asociadas) en el suelo (0-100 cm), con y sin leucaena, 10 años después de su implantación en una pastura megatérmica (Urochloa brizantha-Chloris gayana). Leucaena incrementó el CO, particularmente la forma estable (CO asociado), en un 73% en el horizonte más profundo (50-100 cm). También aumentó el NT en un 12% en la capa superficial (0-20 cm) y en un 21% en el horizonte más profundo. Estos incrementos con leucaena en CO y TN pueden estar asociados a las raíces profundas de leucaena, los aportes adicionales de N por deposición de hojas, por reciclado de nódulos provenientes de la fijación de N y por excretas animales. A pesar del incremento del NT en la superficie, el CO disminuyó en este perfil, lo que se atribuye a la baja cobertura de pasto, consecuencia de una alta presión de pastoreo para controlar la altura de las plantas de leucaena. Bajo un manejo adecuado, leucaena puede mejorar la fertilidad del suelo y la disponibilidad de nitrógeno, además de servir como estrategia de mitigación de gases de efecto invernadero.
Palabras clave: leguminosa arbórea, carbono orgánico del suelo, nitrógeno total.
INTRODUCTION
Grasslands contain approximately 20% of the world’s soil organic carbon (SOC) stocks, indicating their significant role in the global carbon cycle (Puche et al., 2019). However, soils can act as carbon sources due to anthropogenic activities such as intensive livestock grazing, agricultural practices, and other land-use changes that may lead to OC losses. Therefore, improving grassland management for livestock production could be key to enhancing OC sequestration and, consequently, mitigate climate change (Bai and Cotrufo, 2022; Soussana and Menaire, 2014; Lal, 2004).
Grassland management is generally aimed at increasing forage quantity and quality to improve livestock productivity. Nevertheless, improved management practices can also contribute to building soil OC stocks, stimulating plant growth, and protecting OC and nitrogen (TN) in soils (Dondini et al., 2023; Gebremedhn et al., 2022).
Soil carbon and nitrogen interactions play a crucial role in regulating key ecological processes such as nutrient cycling and energy flow (Sardans and Peñuelas, 2012). Adequate nitrogen availability is essential for plant growth, and thus for increasing OC inputs into soils (Dondini et al., 2023). Introducing legumes into grasslands can enhance soil carbon and nitrogen inputs by increasing root biomass, root exudation, and fine root turnover (Bai and Cotrufo, 2022). In this context, the introduction of Leucaena (Leucaena leucocephala ssp. glabrata) into grass pastures could help increase livestock productivity (Radrizzani and Nasca, 2014), enhance soil OC and TN levels (Radrizzani et al., 2011; Conrad et al., 2017; Banegas et al., 2019), and reduce greenhouse gas emissions from livestock (Franzluebbers and Stuedemann, 2009). In previous studies with Leucaena introduction in the region, Banegas et al. (2019) found an increase in OC concentration in the subsoil (20‒100 cm depth), particularly as associated-mineral organic carbon in the deepest horizon (50‒100 cm). The authors also showed an increase in TN concentration, especially in the topsoil, related to an increment of the labile organic nitrogen form.
Soil OC and TN can be fractionated into particulate organic matter (POM) and mineral-associated organic matter (MAOM). These fractions differ in their formation pathways, physical and chemical properties, and mean residence times in the soil. While POM is derived from the fragmentation of plant and microbial residues, MAOM is formed through a process where small organic molecules, leached from plant residues or released from plant roots, interact with minerals directly or indirectly through microbial activity. Microbes can assimilate these small molecules, and their dead cells, called necromass, then become associated with mineral surfaces (Bai and Cotrufo, 2022).
Understanding the response of soil OC, TN, and their respective fractions to grassland management practices is essential for assessing the potential of these soils to sequester atmospheric carbon and the mechanisms that regulate soil carbon cycling (Zimmerman et al., 2006). Due to the limited information available on the integration of tropical tree legumes into pasture systems based on megathermic grasses, we hypothesize that the introduction of Leucaena leucocephala into Chloris gayana cv. Finecut pastures will enhance soil carbon and nitrogen stocks, with a particularly significant contribution at greater soil depths. To test this hypothesis, the study aims to evaluate the quantity and vertical distribution of OC and TN content, as well as their particulate and mineral-associated fractions, in the soil profile (0–100 cm) of a 10-year-old leucaena-grass pasture compared to an adjacent pure tropical grass pasture in the Argentinean Chaco region.
MATERIALS AND METHODS
Site description
This study was conducted at Investigación Animal de la Región Chaco Semiárido (IIACS), operated by the Instituto Nacional de Tecnología Agropecuaria (INTA), located in the western Chaco region (27º11’ S, 65º14’ W; 335 masl), Argentina. The climate is subtropical sub-humid, with a dry season from April to September and an average annual rainfall of 880 mm (75% occurring from October to March). The soil type is classified as a Fluvaquentic Haplustoll (Soil Survey Staff, 1999) (table 1; fig. 2a).
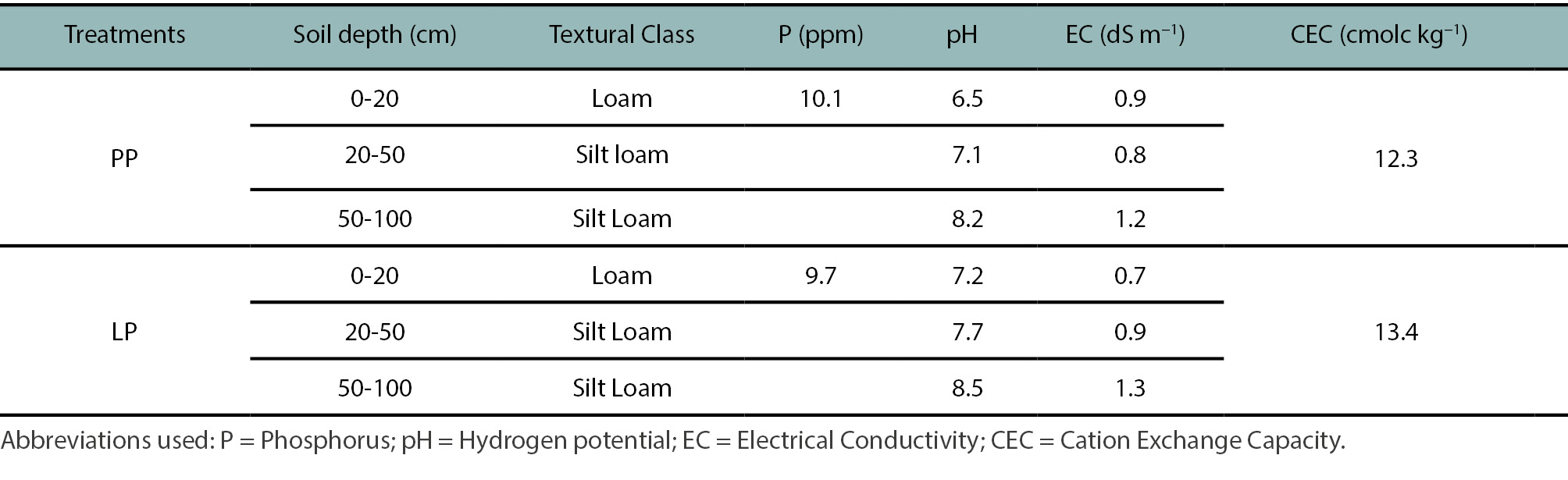
Table 1.
Soil characteristics in Leacaena pasture (LP) and pasture treatment (PP) at Investigación Animal de la Región Chaco Semiárido (IIACS), Tucumán.
Treatments
Four 1-hectare plots were established with a pasture of Urochloa brizantha (syn. Brachiaria brizantha) cv. Marandú (Brachiaria) in 1995. In that homogeneous area, in December 2009, Leucaena cv. K636 was randomly introduced into two of these four plots to evaluate the effect of Leucaena incorporation into ageing pure grass pastures. Leucaena seeds were no-till planted in double-row hedgerows (1 m between rows), with 5 m spacing between adjacent twin hedgerows (figure 1a). In November 2011, the Brachiaria pasture was reseeded with Rhodes grass cv. Finecut (4 kg/ha). Therefore, the treatments evaluated in the present study were as follows: i) Pure pasture (PP): two 1-hectare plots with Urochloa brizantha and Chloris gayana; and ii) Pastures associated with Leucaena leucocephala (LP): two 1-hectare plots with Urochloa brizantha and Chloris gayana accompanying with Leucaena. Both treatments -pure pasture (PP) and Leucaena —enriched pasture (LP)— have been rotationally grazed by Criollo cattle at variable stocking rates, depending on forage availability, from early spring (October) to late autumn (June) (fig. 1b). For most grazing periods, LP was grazed at approximately three times the stocking rate of PP to restrict the height growth of Leucaena, which resulted in overgrazing of the interrow grass (fig. 1d).

Figure 1.
a) Leucaena twin rows zero-till established in December 2009; b) Grazed plots with and without leucaena in February 2011; c) Rhodes grass established in January 2012; d) Low grass cover and high weed presence in March 2019.

Figure 1
a) Leucaena twin rows zero-till established in December 2009; b) Grazed plots with and without leucaena in February 2011; c) Rhodes grass established in January 2012; d) Low grass cover and high weed presence in March 2019.

Figure 1.
a) Leucaena twin rows zero-till established in December 2009; b) Grazed plots with and without leucaena in February 2011; c) Rhodes grass established in January 2012; d) Low grass cover and high weed presence in March 2019.

Figure 1.
a) Leucaena twin rows zero-till established in December 2009; b) Grazed plots with and without leucaena in February 2011; c) Rhodes grass established in January 2012; d) Low grass cover and high weed presence in March 2019.

Figure 2.
a) Initial study of soil properties in September 2011; b) Soil sampling in the pure grass pasture in March 2019.

Figure 2.
a) Initial study of soil properties in September 2011; b) Soil sampling in the pure grass pasture in March 2019.
Soil sampling
Soil samples were collected from four parcels: two with pure pasture (PP) and two with Leucaena-enriched pasture (LP). Sampling took place in both pasture types in March 2019, using twelve 10-meter transects (three per parcel; six per treatment). The collection of soil samples was described in Banegas et al. (2014).
Soil samples were air-dried at 40°C and sieved through a 2 mm mesh. Organic carbon (SOC) concentration was determined using the Walkley–Black method (Nelson and Sommers, 1996), while total nitrogen (TN) concentration was measured using the Kjeldahl method (Bremner, 1965). Fractions of SOC —particulate organic carbon (POC) and mineral-associated organic carbon (MAOC)— and TN —particulate organic nitrogen (PON) and mineral-associated organic nitrogen (MAON)— were quantified following the method described by Cambardella and Elliott (1992). The particulate fraction was obtained by dispersion in 0.5% sodium hexametaphosphate, separated by particle size using sieves ranging from 2000 to 53 µm, and determined as SOC and TN. The associated fraction was determined using the same procedure but working with the fraction smaller than 53 µm (Cambardella and Elliott, 1992).
With these values the ratios C/N, POC/PON, MAOC/MAON, POC/SOC, and MAOC/SOC were also calculated.
To complement the evaluation of the treatments, the stratification ratio (SR) was calculated at depths of 0–20/20–50 cm (SR1) and 0–20/50–100 cm (SR2). This ratio relates the organic carbon (SOC) content of the surface layer to that of the subsurface layer. The rationale for this approach is that the surface layer is strongly influenced by management practices (e.g., tillage, cropping systems, fertilization), whereas the subsurface layer is less affected by such practices (Franzluebbers, 2002). According to this author, the stratification of SOC is a useful index for assessing soil quality, since surface SOC plays a key role in controlling erosion, enhancing infiltration, and conserving nutrients.
To estimate SOC and TN stocks (in tons per hectare), a volumetric conversion was applied using bulk density for the surface soil layer. The SOC and TN contents, expressed as percentages, were then converted into mass values following the approach proposed by Ellert and Bettany (1995).
Grass basal ground cover
Grass basal ground cover (the percentage of the ground covered by plant crowns of each species) was measured in October 2010 and in August 2020 to measure changes over time in botanical composition in relation to PP and LP treatments. Grass basal ground cover was estimated by recording the lengths of intercepts along a eight transects per treatment. Briefly, a 10-m measuring tape was placed on the ground and noting where plant stems or basal parts intersect the tape. The total length of these intercepts was added to calculate the percentage of ground covered by grass species (Urochloa brizantha and Chloris gayana). The total length of the intersections along the 10-m transect represents the proportion (%) of the ground surface occupied by these species.
Statistical analyses
Analysis of variance of soil fertility parameters: SOC, Particulate Organic Carbon (POC), mineral-associated organic carbon (MAOC), TN, Particulate nitrogen (PTN), mineral-associated nitrogen (ATN) and basal grass cover and mean comparisons (Tukey, P<0.05) within pastures were performed to assess the effects of leucaena introduction. Correlation analysis between basal grass cover and SOC was performed using Pearson test. All statistical analyses were carried out using InfoStat software (Di Rienzo et al., 2016).
RESULTS
Soil organic carbon (SOC) and total nitrogen (TN)
Both variables exhibited a clear vertical distribution, with the highest values observed in the 0–20 cm soil layer. At this depth, soil organic carbon (SOC) concentration was higher in PP than in LP (13.8 ± 0.45 g·kg-¹ vs. 12.1 ± 0.75 g·kg-¹ of soil, respectively). In contrast, at the deepest layer (50–100 cm), SOC content was greater in LP compared to PP (6.94 ± 0.22 g·kg-¹ vs. 4.03 ± 0.20 g·kg-¹ of soil, respectively) (fig. 3).
Total nitrogen (TN) concentrations in the topsoil (0–20 cm) were higher in LP than in PP (1.49 ± 0.05 g·kg-¹ vs. 1.33 ± 0.05 g·kg-¹ of soil, respectively). However, no significant differences between treatments were found in the deeper layers (20–50 and 50–100 cm).
Significant differences in the soil C/N ratio were detected at depths of 0–20 cm and 50–100 cm (table 3), with higher values in PP at the surface layer, while LP showed the highest ratio at greater depth.
Organic Carbon and Nitrogen Fractions
Significant differences in POC content were observed at the surface layer (0–20 cm), with higher values in PP (6.77± 0.52 g·kg-¹) than in LP (5.52±0.21 g·kg-¹). Conversely, at deeper layers (50–100 cm), POC was higher in LP (3.44 ± 0.26 g·kg-¹) than in PP (1.81 ± 0.41 g·kg-¹). No significant differences were detected in the intermediate layer. The POC/SOC ratio did not differ significantly across the soil profile. Approximately 49% and 39% of total SOC was associated with the labile fraction in the 0–20 cm and 20–50 cm layers, respectively, under both treatments. At 50–100 cm, this proportion was 49.6% for LP and 44.8% for PP.
MAOC did not differ significantly between treatments in the 0–20 cm layer and 20–50 cm depth (fig. 3). However, at 50–100 cm, MAOC content was higher under LP (3.50 ± 0.38 g·kg-¹) compared to PP (2.22 ± 0.40 g·kg-¹). The MAOC/SOC ratio showed no significant differences between treatments. At 0–20 cm, 51% of SOC was associated with MAOC, while at 20–50 cm, the proportion reached 61% for both treatments. At 50–100 cm, the MAOC/SOC ratio was 50% for LP and 55% for PP.
Nitrogen fractions (PON and MAON) were also vertically stratified across the soil profiles in both treatments. No significant differences in PON content were found between LP and PP at any evaluated depth (fig. 3). In contrast, MAON concentrations were significantly higher in PP than in LP at both 0–20 cm (0.60 ± 0.06 g·kg-¹ vs. 0.43 ± 0.03 g·kg-¹) and 20–50 cm (0.47 ± 0.03 g·kg-¹ vs. 0.30 ± 0.04 g·kg-¹), respectively.
Regarding the carbon-to-nitrogen ratios within fractions —i.e., POC/PON and MAOC/MAON— significant differences between treatments were observed at all depths. In the LP treatment, the POC/PON ratio showed wide variation across the profile, ranging from 6.05 to 20 at deeper layers. In contrast, PP exhibited more stable values, ranging from 6.63 to 9.35. For the MAOC/MAON ratio, higher values were consistently observed in LP compared to PP, ranging from 11 to 14 (table 3).
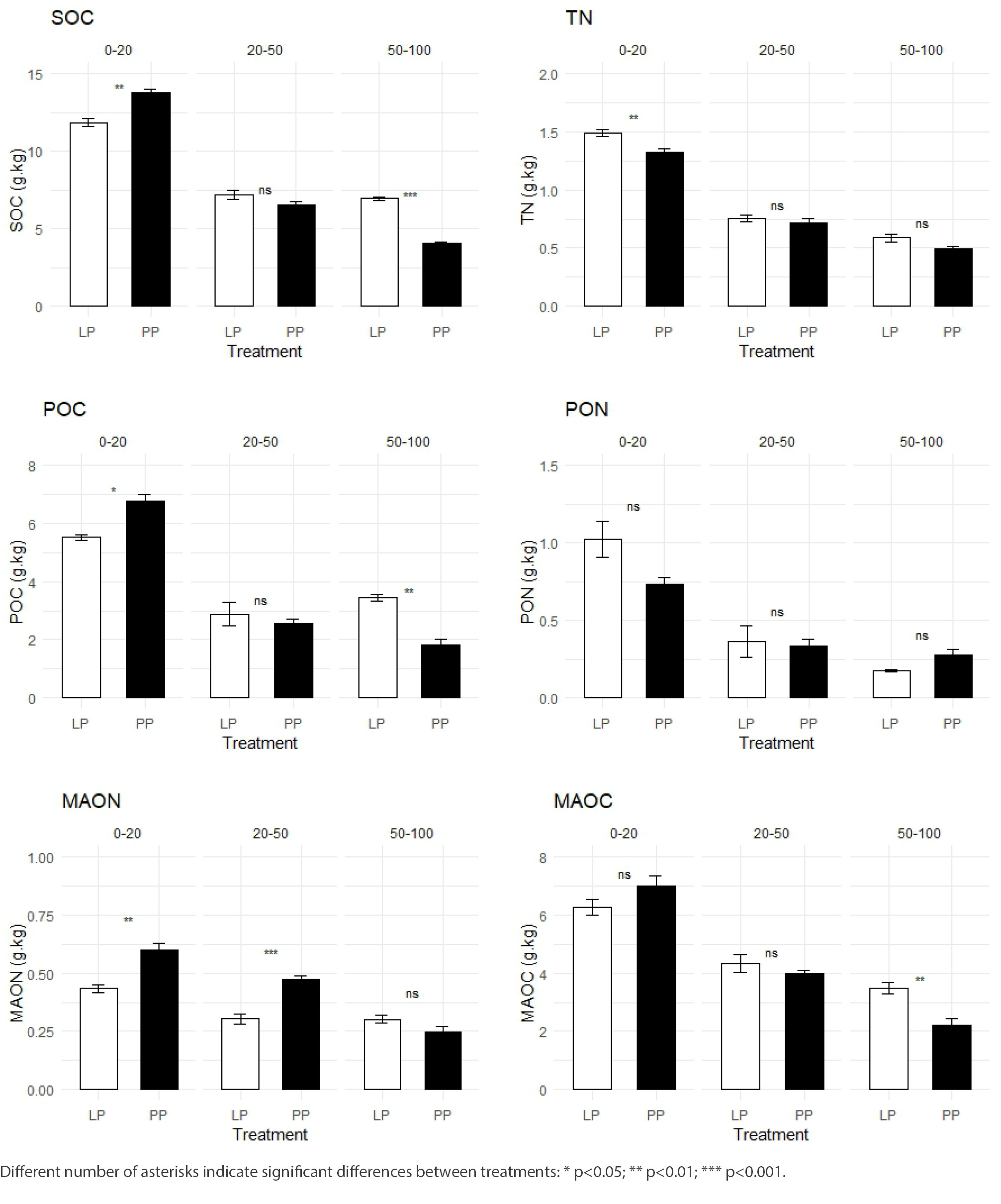
Figure 3.
Concentrations of organic carbon (OC); particulate OC (POC); associate OC (AOC); total nitrogen (TN); particulate organic nitrogen (PON); and associate organic nitrogen (AON), in relation to soil depth (0‒20, 20‒50 and 50‒100 cm horizons) in soils under leucaena-grass pasture (filled squares) and pure grass pasture (open squares) at IIACS-INTA. Means followed by different letters are significantly different (P<0.05); bars represent standard error.
Basal ground cover of pastures
In October 2010, one year after leucaena establishment, grass basal ground cover of sown grass (Urochloa brizantha) was higher in PP than in LP, being 12.73% and 5.67% respectively (p<0.05) (table 1). In August 2020, 9 years after seeding with Chloris gayana, grass basal ground cover of the two sown grasses (Urochloa brizantha and Chloris gayana) was also higher in PP than in LP, being 19.03% and 1.26% respectively (p<0.05) (table 2). In both sampling times, LP was dominated by Cynodon dactylon (18.3% in 2010 and 27.1% in 2020) and broadleaf weeds (3.94% in 2010 and 5.19% in 2020). These results indicate not only significant differences between treatments across both years, but also notable variation between years within each treatment (table 2). A decline in grass condition over time was observed in LP, while an improvement in grass condition was evident in PP.
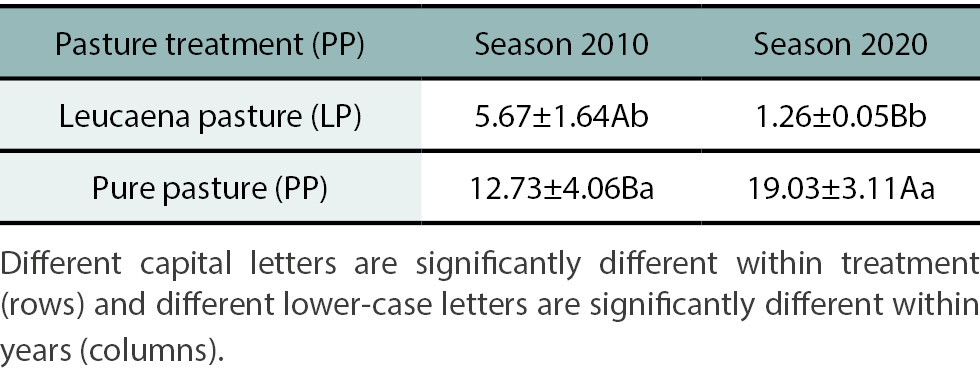
Table 2.
Effect of treatment on grass basal ground cover (%±SE) of sown grasses (Urochloa brizantha in 2010, and Urochloa brizantha and Chloris gayana in 2020) at Investigación Animal de la Región Chaco Semiárido (IIACS) in October 2010 and in August 2020.
DISCUSSION
The vertical distribution of SOC and TN, as well as their fractions in pasture soils, showed similar patterns to those reported in a previous study (Banegas et al., 2019). Although the literature indicates that the inclusion of legumes such as L. leucocephala is expected to improve grass cover and, consequently, increase SOC and TN content in the topsoil (Conrad et al., 2017; Radrizzani et al., 2011), in this study, an increase was observed only in TN. At the soil surface, SOC content was higher under the PP treatment, which may indicate that PP retains more C than N after nine years of livestock production. Moreover, the lower SOC values at 0–20 cm in LP compared to PP suggest that although legumes like L. leucocephala are intended to enhance soil carbon dynamics, the actual outcomes are strongly influenced by management practices and existing grass cover.
This study measured changes over time in basal cover and revealed a clear decline in the condition of sown grasses in LP and an improvement in PP. The decline in grass cover in LP was attributed to the higher stocking rate applied to control the height of Leucaena hedgerows (Radrizzani and Nasca, 2014). Reduced grass cover decreases both litter input and root turnover, thereby limiting Leucaena’s potential contribution to SOC sequestration in the topsoil (Radrizzani et al., 2011). In contrast, the better grass cover observed under the PP treatment enhances litter deposition and root turnover, particularly in the topsoil, where approximately 70% of the root biomass is located within the first 30 cm of the soil profile (Banegas et al., 2020).
The stratification ratio (SR), defined as the ratio of soil organic carbon content in the topsoil to that in the subsoil, has been used as an indicator of SOC sequestration dynamics and soil quality due to its high sensitivity to changes in ecological land types, ecosystems, and management practices (Franzluebbers, 2002). We calculated this ratio for both treatments: SR1 (0–20/20–50 cm) and SR2 (0–20/50–100 cm). In the PP treatment, SR values were 2.12 ± 0.07 for SR1 and 3.43 ± 0.08 for SR2, while in the LP treatment, significantly lower SR values were obtained (1.69 ± 0.12 and 1.75 ± 0.04 for SR1 and SR2, respectively). The lower SR2 value observed in LP supports the hypothesis of reduced organic inputs to the soil surface in this treatment compared to PP, due to a decrease in grass cover and, hence, biomass production.
In both treatments, SR values increased vertically, coinciding with a reduction in SOC content from the topsoil to the subsoil (Zhang et al., 2024). Franzluebbers (2002) states that SOC SR values greater than 2 are rare in degraded conditions, and SR values for soil organic carbon and nitrogen pools above 2 could suggest an improvement in soil quality. These results highlight the importance of promoting adequate grass persistence, emphasizing the need for adaptive management strategies to optimize carbon sequestration benefits in this kind of silvopastoral system (Radrizzani et al., 2019).
SR2 values are also related to the greater SOC content at deeper layers in LP compared to PP. The SOC values observed at the 50–100 cm depth were consistent with those previously reported for these treatments (Banegas et al., 2014). Although various factors can influence SOC accumulation in deeper soil layers, the higher SOC levels under LP in this study were likely due to the deep rooting system of Leucaena, which enhances organic matter inputs at greater depths. These results indicate a sustained SOC presence below 50 cm, which may be attributed to the relative stability of SOC in subsoil layers. At such depths, SOC is less affected by management practices and exhibits slower decomposition rates, owing to the low energy yield of recalcitrant organic compounds and the physical protection provided by stable organo-mineral complexes and soil aggregates (Rolando et al., 2020). Jackson et al. (2017) also mentioned that deep soils contain approximately 68% of soil organic carbon below 30 cm of soil depth.
Like soil OC concentrations, soil TN declined with depth in both pastures, since most of the N (~90%) was bound to SOC in organic matter. These results are consistent with previous studies, which reported higher total nitrogen (TN) concentrations in the topsoil (0‒15 cm) of Leucaena–grass silvopastoral systems compared to grass-only systems (Conrad et al., 2017, 2018; Radrizzani et al., 2011). In similar analyses, differences between treatments were observed even at greater depths (up to 1 meter). However, in this study, no significant differences between treatments were found below the 0–20 cm layer. Differences between treatments were observed mainly in the topsoil and could be attributed to both the recycling of N-rich residues and biological N fixation after legume introduction. In concordance, Imogie et al. (2008) remarked that the enhancement of soil nutrient levels can be credited to L. leucocephala, which, alongside its nitrogen-fixing properties, plays a significant role in enriching the soil through the decomposition of its fallen leaves and in increasing nitrogen and organic matter content in the soil. The same authors suggest that L. leucocephala also promotes a favorable microclimate that stimulates soil microfauna activity, which, in turn, improves the soil’s water retention capacity and promotes nutrient recycling.
The C:N ratios ranged from 8.14 to 11.91 (table 3). The values obtained at different depths for the treatments reflects the dynamics of SOC and TN accumulation in each system, with greater carbon input at depth in PP and higher nitrogen input at the surface in LP.
Separating soil organic matter into particulate organic matter (POM) and mineral-associated organic matter (MAOM) is funda mental for understanding SOM cycling (Villarino et al., 2023). The labile soil organic matter fractions, such as particulate organic carbon and nitrogen, are characterized by their rapid turnover and are considered sensitive indicators of the effects of management practices (Yang et al., 2018). The associated organic matter fraction (in this case, MAOC and MAON) is considered a stable fraction. It is mainly comprised by labile compounds (i.e., low molecular weight) that are linked to mineral soil particles (clay and silt) and protected against decomposition (Cotrufo et al., 2013). So, management practices that stimulate their exposition to microbial activity, can also promote their quickly decomposition (Santos et al., 2020).
Our results show that the distribution of these fractions across the systems was differential. The higher surface POC values (0-20 cm) in PP are consistent with previous studies reporting elevated levels of this labile fraction in grasslands suggests that the accumulation is further promoted by conservation management practices that minimize soil disturbance (Semenov et al., 2019; Franzluebbers, 2024a). In a similar vein, several authors highlight that grassland management involving perennial forage cover qualifies as a conservation land use due to its positive effects on enhancing POC, mainly related to greater biomass input and SOM is not yet completely stabilized (Rodruigues de Oliveira et al., 2022). Conversely, grasslands subjected to poor management practices —characterized by reduced income due to lower aboveground productivity, lower quality, or both— may pose risks to environmental quality, including the degradation of the labile fraction of organic matter, as observed in LP (Franzluebbers, 2024a).
Subsoil carbon fractions have received little attention, as levels are often assumed to stabilize below 30 cm (Rodrigues Oliveira et al., 2022; Schmidt et al., 2011). At deepest layer, POC and MAOC content can be altered by tillage and subsoiling, root distribution, fertilization and burning that induce priming, and erosion exposing deeper layers (Rodrigues Oliveira et al., 2022). In this study, vegetation type, specifically the inclusion of a shrub–tree species such as Leucaena in LP treatment with a deeper root system, may have accounted for the observed differences at 50-100 cm of soil depth. These results highlight that the incorporation of deep-rooted legumes enhances SOC by simultaneously increasing both fractions, mainly through root biomass and exudates, which are now recognized as efficient contributors to SOC (Villarino et al., 2021).
Although N-POM content in the different soil layers was not particularly effective in distinguishing among the evaluated systems, the distribution of this fraction in the surface layer revealed a particular behavior between treatments. In LP, N-POM represented 69% of total nitrogen (TN), whereas in PP it accounted for only 56%. These results indicate that the relative contribution of N-POM to TN in the topsoil is closely linked to the type of management system. In particular, the incorporation of a leguminous species enhanced the buildup of organic nitrogen, mainly through its allocation into the more labile and readily available fraction. This is also observed in the C/N ratio of this fraction, where the lowest values in the upper soil layers were found in LP, showing a marked N enrichment in the labile fraction of this system (table 3).
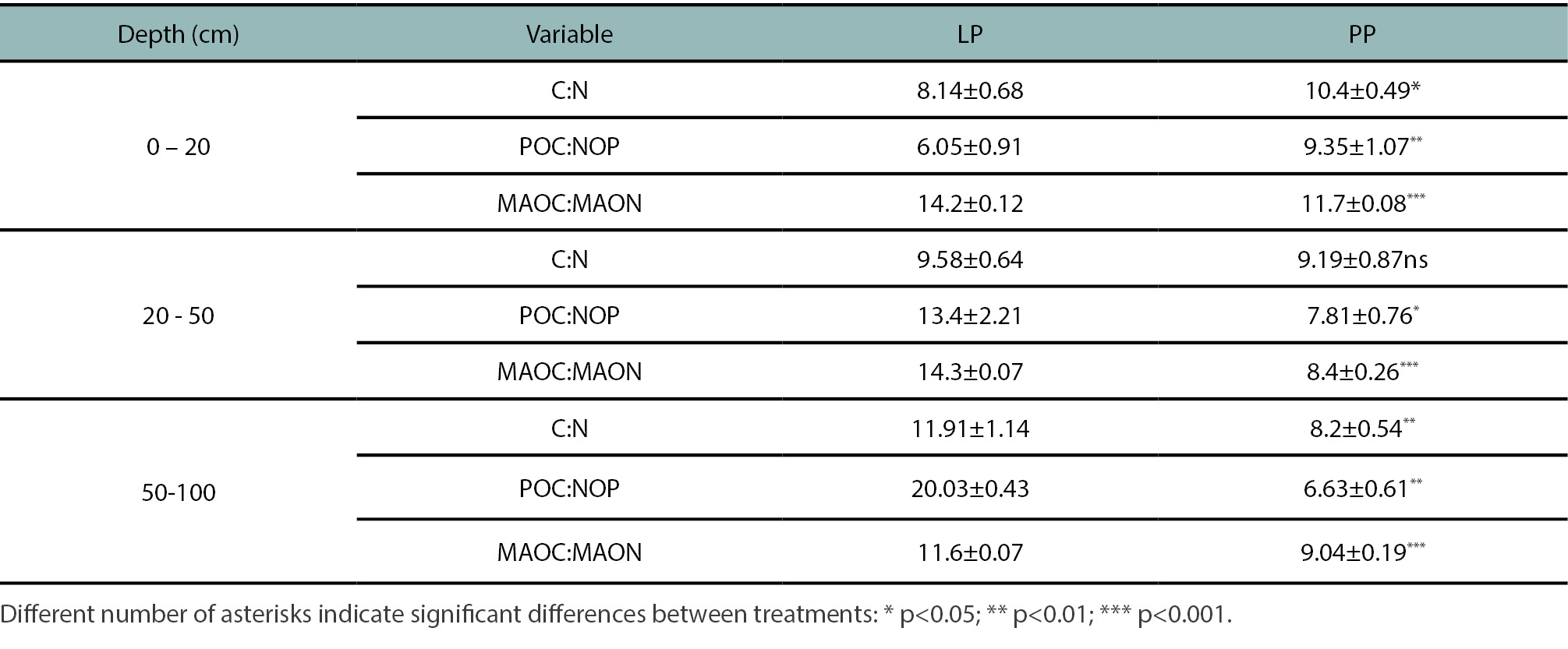
Table 3.
Average C:N ratios for soil (OC:TN) and organic fractions (POC:NOP, MAOC:MAON) in Leucaena pasture (LP) and pasture treatment (PP).
In contrast, N in the stable fraction was higher in PP than in PL. Although residue composition (C:N ratio) was not analyzed in this study, Manzoni and Cotrufo (2024) indicate that residues relatively poor in N exhibit strong immobilization and accumulation of N in MAON, due to the preferential retention and stabilization of this N during the decomposition of residues with a high C:N ratio
For the same soil mass (2640 Mg/ha), PP stored higher SOC than LP (36.43 Mg SOC/ha vs. 32 Mg SOC/ha, respectively). In comparison to the previous soil survey, done in the same pastures 5 years after leucaena establishment (2014) (Banegas et al., 2019), SOC stocks increased by 1.77 Mg/ha under PP, reaching an annual increment rate of 0.35 Mg/ha/year, whereas the level of SOC in PL remained stable. The annual increment rate in PP is consistent with other results observed by Conant et al. (2017), who reported an average SOC increment of 0.47 Mg/ha/year in grazed grass pastures. In contrast to SOC, the increment of TN stock during the same period was higher in LP than in PP (0.10 Mg/ha vs. 0.07 Mg/ha), leading to an increment rate of 0.020 and 0.014 Mg/ha/year for LP and PP, respectively. The rate of TN increase under LP is consistent with the values reported by Radrizzani et al. (2011) in Australia, in a 20, 31 and 38-year-old leucaena-grass pastures (0.017, 0.011 and 0.014 Mg/ha/year, respectively). This focuses that while leucaena integration promotes TN accumulation, it may not lead to SOC sequestration without tailored grazing strategies.
CONCLUSIONS
This study evaluated the vertical distribution of soil organic car bon (SOC) and total nitrogen (TN), as well as their particulate and mineral-associated fractions, in soils under a 10-year-old Leucaena leucocephala–grass silvopastoral system and an adjacent grass-only pasture in the Argentinean Chaco region. The results revealed distinct patterns of C and N allocation between systems and across soil depths.
While the grass-only pasture (PP) accumulated higher SOC in the surface layers, the leucaena-based pasture (LP) showed higher TN contents, particularly within the labile fraction. These findings suggest that the integration of Leucaena enhances nitrogen enrichment through biological fixation and residue input but does not necessarily increase surface SOC without proper grazing management. Conversely, greater SOC contents at deeper soil layers under LP indicate that deep-rooted legumes contribute to subsoil carbon stabilization.
This study highlights the importance of integrating adaptive management strategies in silvopastoral systems to optimize both carbon sequestration and nitrogen cycling, reinforcing their potential as sustainable land-use alternatives in the Chaco region.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported by Instituto de Investigación Animal de la Región Chaco Semiárido (IIACS), operated by the Instituto Nacional de Tecnología Agropecuaria (INTA).
REFERENCES
BAI, Y.; COTRUFO, M.F. 2022. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science, 377(6606), 603-608. https://doi.org/10.1126/science.abo2380
BANEGAS, N.; CORBELLA, R.; VIRUEL, E.; PLASENCIA, A.; ROIG, B.; RADRIZZANI, A. 2019. Leucaena leucocephala introduction into a tropical pasture in the Chaco region of Argentina. Effects on soil carbon and total nitrogen. Tropical Grasslands-Forrajes Tropicales, 7, 295-302. https://doi.org/10.17138/TGFT(7)295-302
BANEGAS, N.; DOS SANTOS, D.; GUERRERO MOLINA, F.; ALBANESI, A. 2020. Glomalin contribution to soil organic carbon under different pasture management in a saline soil environment. Archives of Agronomy and Soil Science, 68(3), 1-15.
BONDARUK, V.; LEZAMA, F.; DEL PINO, A.; PIÑEIRO, G. 2020. Overseeding legumes in natural grasslands: Impacts on root biomass and soil organic matter of commercial farms. Science of the Total Environment, 74. https://doi.org/10.1016/j.scitotenv.2020.140771
BREMNER, J.M. 1965. Total nitrogen. In: NORMAN, A.G. (Ed.). Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties. Agronomy Monograph 9.2. ASA and SSSA. 595-624 pp.
CAMBARDELLA, C.A.; ELLIOTT, E.T. 1992. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Science Society of America Journal, 56, 777-783. https://doi.org/10.2136/sssaj1992.03615995005600030017x
CONANT, R.; CERRI, C.; OSBORNE, B.; PAUSTIAN, K. 2017. Grassland management impacts on soil carbon stocks: a new synthesis. Ecological Applications, 27(2), 662-668.
CONRAD, K.A.; DALAL, R.C.; DALZELL, S.A.; ALLEN, D.E.; MENZIES, N.W. 2017. The sequestration and turnover of soil organic carbon in subtropical Leucaena-grass pastures. Agriculture, Ecosystems & Environment, 248, 38-47. https://doi.org/10.1016/j.agee.2017.07.020
COTRUFO, M.F.; WALLENSTEIN, M.D.; BOOT, C.M.; DENEF, K.; PAUL, E. 2013. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Global Change Biology, 19(4), 988-995. https://doi.org/10.1111/gcb.12113
DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZÁLEZ, L.; TABLADA, M.; ROBLEDO, C.W. 2016. InfoStat, Version 2016. Grupo InfoStat, F.C.A., Universidad Nacional de Córdoba, Argentina. www.infostat.com.ar
DONDINI, M.; MARTIN, M.; CAMILLIS, C.; UWIZEYE, A.; SOUSSANA, J.F.; ROBINSON, T.; STEINFELD, H. 2023. Global assessment of soil carbon in grasslands: from current stock estimates to sequestration potential. FAO, Rome.
FISHER, M.J.; RAO, I.M.; AYARZA, M.A.; LASCANO, C.E.; SANZ, J.I.; THOMAS, R.J.; VERA, R.R. 1994. Carbon storage by introduced deep-rooted grasses in the South America savannas. Nature, 371, 236-238. https://doi.org/10.1038/371236a0
FERREIRA, E.; ALVES, B.J.R.; URQUIAGA, S.; BODDEY, R.M. 1995. Perdas de N derivado das fezes bovinas depositadas na superficie do solo. Proceedings of the xxxii Reuniao Anual da Sociedade Brasileira de Zootecnia, Brasilia, Brazil. 125-126 pp.
FOLLETT, R.F.; LEAVITT, S.W.; KIMBALL, J.M.; PRUESSNER, E.G. 2003. Paleo-environmental inferences from δ13C of soil organic C in 14C-dated profiles in the U.S. Great Plains. xvi INQUA Congress, Reno, NV, USA.
FRANZLUEBBERS, A.J. 2002. Soil organic matter stratification ratio as an indicator of soil quality. Soil and Tillage Research, 66(2), 95-106.
FRANZLUEBBERS, A.J.; STUEDEMANN, J.A. 2009. Soil-profile organic carbon and total nitrogen during 12 years of pasture management in the Southern Piedmont USA. Agriculture, Ecosystems & Environment, 129, 28-36. https://doi.org/10.1016/j.agee.2008.06.013
GARTEN, C.T.; WULLSCHLEGER, S.D. 2000. Soil carbon dynamics beneath switchgrass as indicated by stable isotope analysis. Journal of Environmental Quality, 29, 645-653.
GEBREMEDHN, H.H.; KELKAY, T.Z.; TESFAY, Y.; TUFFA, S.; DEJENE, S.W.; MENSAH, S.; DEVENISH, A.J.M.; EGERU, A. 2022. Carbon Stock and Change Rate under Different Grazing Management Practices in Semiarid Pastoral Ecosystem of Eastern Ethiopia. Land, 11, 639. https://doi.org/10.3390/land11050639
IMOGIE, A.E.; UDOSEN, C.V.; UGBAH, M.M.; UTULU, S.N. 2008. Long term effect of Leucaena leucocephala on soil physico-chemical properties and fresh fruit bunch (FFB) production of oil palm. African Journal of Plant Science, 2(11), 129-132.
JACKSON, R.B.; LAJTHA, K.; CROW, S.E.; HUGELIUS, G.; KRAMER, M.G.; PIÑEIRO, G. 2017. The ecology of soil carbon: pools, vulnerabilities, and biotic and abiotic controls. Annual Review of Ecology, Evolution, and Systematics, 48, 419-445.
JOBBÁGY, E.G.; JACKSON, R.B. 2000. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 10(2), 423-436.
LAL, R. 2003. Soil erosion and the global carbon budget. Environmental International, 29, 437-450.
LAL, R. 2004. Agricultural activities and the global carbon cycle. Nutrient Cycling in Agroecosystems, 70, 103-116.
LASCANO, C.E.; PETERS, M. 2004. Forage quality and utilization. Forage Seed Production and Quality. CABI Publishing. 151-165 pp.
LIU, Y.; CHEN, L.; DUAN, W.; BAI, Y.; LI, X. 2022. Effects of litter decomposition on soil N in Picea mongolica forest at different forest ages. Forests, 13(4), 520. https://doi.org/10.3390/f13040520
LODGE, G.M.; KING, K.L.; HARDEN, S. 2006. Effects of pasture treatments on detached pasture litter mass, quality, litter loss, decomposition rates, and residence time in northern New South Wales. Australian Journal of Agricultural Research, 57, 1073-1085. https://doi.org/10.1071/AR05408
NELSON, D.; SOMMERS, L.E. 1996. Total carbon, organic carbon, and organic matter. In: SPARKS, D.L.; PAGE, A.L.; HELMKE, P.A.; LOEPPERT, R.H. (Eds.). Methods of Soil Analysis, Part 3, Chemical Methods. ASA and SSSA. 961-1010.
PACHAS, A.N.A.; SHELTON, H.M.; LAMBRIDES, C.J.; DALZELL, S.A.; MURTAGH, G.J.; HARDNER, C.M. 2018. Effect of tree density on competition between Leucaena leucocephala and Chloris gayana using a Nelder Wheel trial. ii. Belowground interactions. Crop and Pasture Science, 69(7), 733-744.
PANDEY, C.B.; SHARMA, D.K.; BARGALI, S.S. 2006. Decomposition and nitrogen release from Leucaena leucocephala in central India. Tropical Ecology, 47(1), 149-152.
PUCHE, N.; SENAPATI, N.; FLECHARD, C.R.; KLUMPP, K.; KIRSCHBAUM, M.U.F.; CHABBI, A. 2019. Modeling carbon and water fluxes of managed grasslands: Comparing flux variability and net carbon budgets between grazed and mowed systems. Agronomy, 9(4), 183. https://doi.org/10.3390/agronomy9040183
RADRIZZANI, A. 2009. Long-term productivity of leucaena (Leucaena leucocephala)-grass pastures in Queensland. [Ph.D. Thesis. The University of Queensland, Brisbane, Australia]. espace.library.uq.edu.au/view/UQ:195473
RADRIZZANI, A.; SHELTON, H.M.; DALZELL, S.A.; KIRCHHOF, G. 2011. Soil organic carbon and total nitrogen under Leucaena leucocephala pastures in Queensland. Crop and Pasture Science, 62, 337-345. https://doi.org/10.1071/CP10115
RADRIZZANI, A.; NASCA, J.A. 2014. The effect of Leucaena leucocephala on beef production and its toxicity in the Chaco Region of Argentina. TROPICAL GRASSLANDS-FORRAJES TROPICALES, 2, 127-129. https://doi.org/10.17138/tgft(2)127-129
RADRIZZANI, A.; PACHAS, A.N.A.; GÁNDARA, L.; NENNING, F.; PUEYO, D. 2019. Leucaena feeding systems in Argentina. ii. Current uses and future research priorities. Tropical Grasslands-Forrajes Tropicales, 7, 389-396. https://doi.org/10.17138/TGFT(7)389-396
SANTOS, R.S.; OLIVEIRA, F.C.; FERREIRA, G.W.; FERREIRA, M.A.; ARAÚJO, E.F.; SILVA, I.R. 2020. Carbon and nitrogen dynamics in soil organic matter fractions following eucalypt afforestation in southern Brazilian grasslands (Pampas). Agriculture, Ecosystems & Environment, 301, 106979. https://doi.org/10.1016/j.agee.2020.106979
SARDANS, J.; RIVAS-UBACH, A.; PEÑUELAS, J. 2012. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspectives in Plant Ecology, Evolution and Systematics, 14(1), 33-47. https://doi.org/10.1016/j.ppees.2011.08.002
SHELTON, H.M.; FRANZEL, S.; PETERS, M. 2005. Adoption of tropical legume technology around the world: Analysis of success. Tropical Grasslands, 39(4), 216-228.
SOIL SURVEY STAFF. 1999. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys. 2nd Edn. Natural Resources Conservation Service, USA. Department of Agriculture Handbook 436.
SOUSSANA, J.F.; MENAIRE, G. 2014. Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems. Agriculture, Ecosystems & Environment, 190, 9-17.
WANDER, M. 2004. Soil organic matter fractions and their relevance to soil function. Soil Organic Matter in Sustainable Agriculture. CRC Press, Boca Raton, FL. 67-102 pp.
YANG, F.; TIAN, J.; MEERSMANS, J.; FANG, H.; YANG, H.; LOU, Y.; LI, Z.; LIU, K.; ZHOU, Y.; BLAGODATSKAYA, E.; KUZYAKOV, Y. 2018. Functional soil organic matter fractions in response to long-term fertilization in upland and paddy systems in South China. CATENA, 162, 270-277. https://doi.org/10.1016/j.catena.2017.11.004
YONI, M.; SAKO, A.; ABBADIE, L.; SERPANTIÉ, G. 2018. Carbon and nitrogen dynamics of soil organic matter fractions in thickets and intergrowth areas of Sudanian savannah grasslands, Bondoukuy, Western Burkina Faso. Environment and Natural Resources Research, 8(2), 1-14.
YOUKHANA, A.; IDOL, T. 2009. Tree pruning mulch increases soil C and N in a shaded coffee agroecosystem in Hawaii. Soil Biology and Biochemistry, 41(12), 2527-2534. https://doi.org/10.1016/j.soilbio.2009.09.011
ZHANG, Q.; YUE, C.; YU, P.; XU, H.; WU, J.; SHENG, F. 2024. Soil organic carbon storage and stratification in land use types in the source area of the Tarim River Basin. Sustainability, 16(24), 11255. https://doi.org/10.3390/su162411255
ZIMMERMANN, M.; LEIFELD, J.; SCHMIDT, M.; SMITH, P.; FUHRER, J. 2006. Measured soil organic matter fractions can be related to pools in the RothC model. European Journal of Soil Science, 58(3), 658-667.
Información adicional
redalyc-journal-id: 864