Abstract: Melatonin (N-acetyl-5-methoxytryptamine) has emerged as a key bioactive molecule in plants, extending far beyond its well-known role in animals. In horticultural crops, melatonin contributes to physiological regulation, stress mitigation, and improved agronomic performance. This review synthesizes current evidence on melatonin’s biosynthesis, physiological roles, and its effects on tolerance to abiotic stressors such as drought, salinity, and temperature extremes. The review further discusses its application in species including lettuce, tomato, cucumber, and broccoli, with a focus on seed treatment, foliar application, and irrigation. Despite promising outcomes, challenges remain in standardizing doses, validating field efficacy and integrating melatonin into commercial horticulture. Future directions highlight the need for omics-based studies, field-level validations and exploration of melatonin-based technologies to support sustainable crop production.
Keywords: melatonin, abiotic stress mitigation, antioxidant defense, crop productivity, hormonal regulation.
Resumen: La melatonina (N-acetil-5-metoxitriptamina) ha emergido como una molécula bioactiva clave en las plantas, extendiéndose mucho más allá de su conocido papel en los animales. En los cultivos hortícolas, la melatonina contribuye a la regulación fisiológica, la mitigación del estrés y la mejora del rendimiento agronómico. Esta revisión sintetiza la evidencia actual sobre la biosíntesis de la melatonina, sus roles fisiológicos y sus efectos en la tolerancia a estresores abióticos como la sequía, la salinidad y las temperaturas extremas. La revisión aborda además su aplicación en especies como la lechuga, el tomate, el pepino y el brócoli, centrándose en el tratamiento de semillas, la aplicación foliar y la irrigación. A pesar de que los resultados son prometedores, persisten desafíos en la estandarización de las dosis, la validación de la eficacia en campo y la integración de la melatonina en la horticultura comercial. Las direcciones futuras resaltan la necesidad de estudios basados en ómicas, validaciones a nivel de campo y la exploración de tecnologías basadas en melatonina para apoyar la producción de cultivos sostenible.
Palabras clave: melatonina, mitigación del estrés abiótico, defensa antioxidante, productividad de cultivos, regulación hormonal.
Melatonin in horticultural crops: roles in stress tolerance and agronomic applications
Instituto Nacional de Tecnología Agropecuaria

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Horticultural crops, which include vegetables, fruits and ornamental species, are characterized by their high commercial value, perishability and sensitivity to environmental conditions. Unlike many staple crops, horticultural species often have shallow root systems, rapid growth cycles and delicate physiological balances, making them particularly vulnerable to abiotic stresses such as drought, salinity, extreme temperatures, heavy metal toxicity, and ultraviolet (UV) radiation (Rouphael and Colla, 2020). These stressors disrupt vital cellular processes, leading to oxidative damage, impaired photosynthesis, altered hormonal signaling and reduced nutrient uptake (Ahanger et al., 2017; Hasanuzzaman et al., 2013). In crops like tomato (Solanum lycopersicum L.), lettuce (Lactuca sativa L.), cucumber (Cucumis sativus L.) and pepper (Capsicum annuum L.), even moderate levels of abiotic stress can cause drastic reductions in yield, quality, shelf life, and visual appeal—factors that are critical in competitive markets and for food chain sustainability (Francis et al., 2012). Additionally, the intensification of climate variability and the expansion of production into marginal soils further increase the exposure of horticultural systems to abiotic challenges (Borgohain et al., 2019), highlighting the urgent need for effective and sustainable stress mitigation strategies.
Melatonin (N-acetyl-5-methoxytryptamine) was first discovered in animals (Lerner et al., 1958), where it plays a central role in regulating circadian rhythms, sleep-wake cycles, reproductive timing and antioxidant defenses (Karasek and Winczyk, 2006). In humans, melatonin is primarily synthesized in the pineal gland and is widely used as a natural supplement for sleep disorders, jet lag and neurodegenerative conditions due to its ability to scavenge free radicals and modulate immune responses (Reiter et al., 2014).
More recently, the presence and synthesis of melatonin have been confirmed in a wide range of plant species (Dubbels et al., 1995; Hattori et al., 1995), where it is now recognized as a pleiotropic molecule involved in numerous physiological and developmental processes. In plants, melatonin contributes to seed germination (Wang et al., 2024), root architecture (Pelagio‐Flores et al., 2012), leaf senescence delay (Liang et al., 2015), photosynthetic efficiency (Wang et al., 2016) and fruit ripening (Liu et al., 2016). Importantly, its powerful antioxidant activity (Oloumi, 2022), capacity to regulate gene expression (Fan et al., 2018), interaction with plant hormones such as auxins and abscisic acid (ABA) (Arnao and Hernández-Ruiz, 2018) and its involvement in mitochondrial protection (Turk and Genisel, 2019) make it a promising candidate for improving abiotic stress tolerance. These multifaceted roles have sparked growing interest in using melatonin as a natural, sustainable biostimulant in horticultural crop management (Bose and Howlader, 2020).
Melatonin (N-acetyl-5-methoxytryptamine) is endogenously synthesized in plants through a conserved pathway that begins with the amino acid tryptophan. This precursor is sequentially converted into tryptamine by tryptophan decarboxylase (TDC), then, into serotonin via tryptamine 5-hydroxylase (T5H). Finally, serotonin is acetylated by serotonin N-acetyltransferase (SNAT) and methylated by acetylserotonin O-methyltransferase (ASMT) or caffeic acid O-methyltransferase (COMT), leading to melatonin production (fig. 1). This multi-step route is highly flexible and may vary depending on tissue type, developmental stage, and environmental conditions (Back et al., 2016).
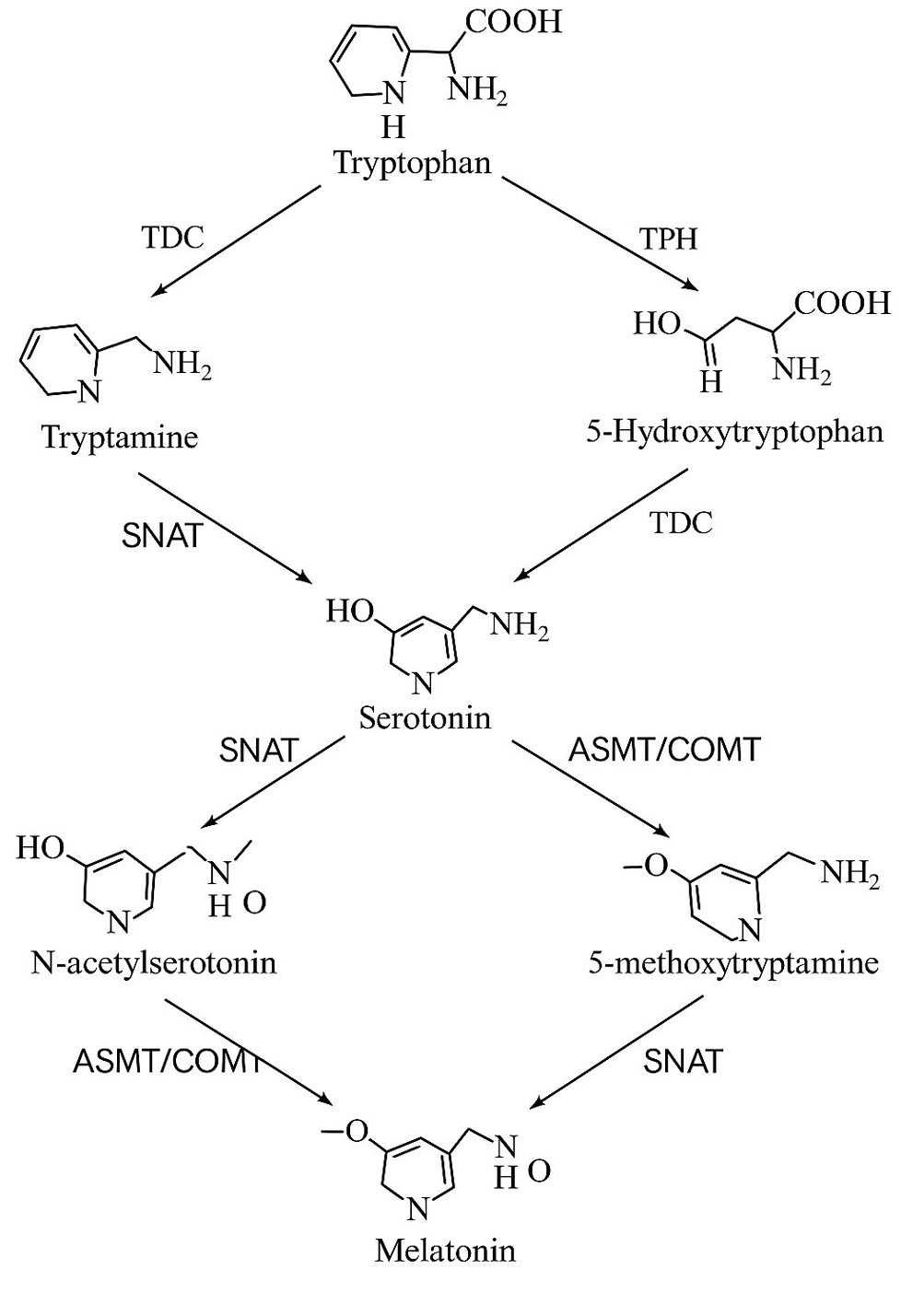
Figure 1.
Melatonin biosynthesis pathway in plants (adapted from Yan et al., 2023).
Unlike animals, where melatonin is primarily synthesized in the pineal gland, plants produce melatonin in multiple subcellular compartments, including the cytoplasm, chloroplasts, mitochondria, and even the nucleus. This widespread biosynthesis allows melatonin to act both locally and systemically, supporting its role as a versatile regulator in plant physiology (Zhao et al., 2019).
Melatonin distribution varies widely across tissues and species. It has been detected in roots, leaves, flowers, fruits, and seeds, with concentrations fluctuating in response to developmental signals and external stimuli, such as light, temperature, and oxidative stress. Notably, higher melatonin levels are often found in organs with elevated metabolic activity or exposure to stress, which suggests a protective allocation mechanism within the plant (Arnao and Hernández Ruiz, 2015).
In this context, melatonin is not merely a byproduct of tryptophan metabolism, but an actively regulated molecule with specific functions. Its spatial distribution across plant organs and cellular compartments points to a complex role that extends far beyond antioxidant activity, positioning melatonin as an integral component of the plant’s adaptive response network.
PHYSIOLOGICAL ROLES OF MELATONIN IN PLANTS
Melatonin has gained increasing attention as a multifunctional regulator in plants, including horticultural crops such as fruits, vegetables, and ornamentals. Beyond its well-established antioxidant properties, melatonin plays critical roles in modulating growth, development, photosynthesis, reproductive processes, and secondary metabolism under normal physiological conditions (Arnao, 2016; Fan et al., 2018). Due to its broad spectrum of actions, melatonin serves as a central signaling molecule in plant biology.
Regulation of plant growth and development
Melatonin acts as a multifunctional growth regulator in horticultural crops, influencing key stages of plant development from seed germination to the formation of reproductive organs. Its ability to interact with the plant hormonal network —particularly auxins, gibberellins, and cytokinins— allows it to coordinate complex growth responses in a dose- and context-dependent manner (Kołodziejczyk and Posmyk, 2016). These interactions suggest that melatonin is not acting independently, but rather as a component of a tightly regulated signaling web that orchestrates morphogenesis and developmental plasticity.
In root development, melatonin has shown auxin-like activity, promoting lateral root formation, root elongation, and root biomass accumulation. Studies in tomato, cucumber, and pea (Pisum sativum L.) have demonstrated that exogenous melatonin at low micromolar concentrations enhances root system architecture, likely through the modulation of auxin transport and signaling pathways (Tian et al., 2024; Wang et al., 2022; Yusuf et al., 2020). These improvements in root morphology enhance water and nutrient uptake efficiency, which are critical for optimal growth and yield.
During vegetative growth, melatonin contributes to shoot elongation and leaf expansion by modulating cell division and cell wall loosening genes (Wang et al., 2022). In broccoli (Brassicaoleracea L.), melatonin treatment under controlled conditions resulted in improved leaf size and fresh weight accumulation (Sardar et al., 2023), highlighting its potential as a growth-promoting agent. In root development, studies in cucumber demonstrated that melatonin enhances lateral root formation and increases root biomass (Li et al., 2025), which is essential for nutrient absorption and anchorage. Moreover, melatonin-treated plants typically exhibit improved biomass accumulation and chloroplast development during early growth stages, contributing to vigorous seedling establishment (Sharif et al., 2018; Zhao et al., 2021).
Melatonin also delays leaf senescence by regulating ROS homeostasis and modulating senescence-associated gene expression (Oloumi, 2022). This delay in aging supports prolonged photosynthetic activity and resource allocation to developing tissues. Additionally, melatonin enhances the accumulation of primary metabolites, including soluble sugars and proteins, contributing to better nutritional quality and plant vigor (Sharif et al., 2018).
Taken together, these findings underscore the role of melatonin as a fine-tuner of vegetative growth in horticultural crops, acting through a combination of antioxidant protection, hormonal crosstalk, and gene regulatory mechanisms. These effects not only improve growth under optimal conditions but also prepare the plant for enhanced resilience to future stress, which is further explored in subsequent sections.
Role in reproductive development and fruit quality
Regarding reproductive development, melatonin has been linked to enhanced floral initiation and reduced floral abscission (Arnao and Hernández-Ruiz, 2020), although this area remains underexplored in field-grown horticultural crops. Preliminary evidence from ornamental species suggests that melatonin may support flower retention by stabilizing hormonal balance and improving antioxidant capacity in floral tissues (Hosseini et al., 2025; SeyedHajizadeh et al., 2024; Wang et al., 2024).
In ‘Micro-Tom’, a model cultivar of tomato, melatonin accumulation was observed throughout fruit development, suggesting a regulatory role for melatonin in the physiological processes associated with fruit maturation (Okazaki and Ezura, 2009).
In strawberry (Fragaria × ananassa Duch.), the exogenous application of 10 µM melatonin delayed fruit ripening by lowering ABA and H₂O₂ levels, enhancing antioxidant activity, and altering key biochemical and morphological characteristics. These findings suggest that melatonin modulates the ripening process through hormonal regulation and ROS-related mechanisms (Mansouri et al., 2023).
Furthermore, melatonin improves fruit quality attributes including sugar accumulation, firmness, pigment content and aroma compound synthesis (Arnao and Hernández-Ruiz, 2020). In strawberries, for instance, preharvest melatonin application increased soluble sugar content and enhanced anthocyanin accumulation, leading to improved coloration and sweetness (El-Mogy et al., 2019).
Seed physiology and germination
Melatonin plays a crucial role in seed physiology, functioning both as a signaling molecule and a protective antioxidant during seed maturation and germination. Of melatonin have been detected in seeds of several edible horticultural species (Manchester et al., 2000), indicating its potential role in safeguarding seeds during dormancy. In mature seeds, which are typically dry and metabolically inactive, melatonin likely serves as an intrinsic antioxidant reservoir. Its accumulation is thought to offer preemptive protection against oxidative damage during storage and the initial phases of imbibition, when enzymatic antioxidant systems are not yet fully active (Korkmaz et al., 2014).
During germination, melatonin contributes to improved seed performance by enhancing water uptake, activating hydrolytic enzymes such as amylases and proteases, and mitigating oxidative stress through the reduction of lipid peroxidation and reactive oxygen species (ROS) accumulation (Wang et al., 2022). These physiological improvements lead to higher germination rates, more uniform seedling emergence, and enhanced early vigor. In horticultural crops like lentils (Lens culinaris L.), bean (Phaseolus vulgaris L.), and cucumber exogenous application of melatonin at low micromolar concentrations has been shown to accelerate germination under both optimal and suboptimal conditions (Aguilera et al., 2015; Posmyk et al., 2009). However, these effects are dose-dependent, and excessive melatonin levels can inhibit germination or delay radicle emergence (Castañares and Bouzo, 2019).
Beyond its antioxidant properties, melatonin is also involved in the hormonal regulation of germination. It appears to favor a pro-germinative hormonal balance by upregulating gibberellin (GA) biosynthesis while downregulating ABA signaling (Chen et al., 2021).
Overall, melatonin emerges as a promising biostimulant to enhance seed germination and seedling establishment in horticultural crops, particularly under stress-prone conditions, such as salinity, drought or extreme temperatures, topics that will be further explored in the following chapter.
Circadian and photoperiodic regulation by melatonin
Melatonin is widely recognized as a key regulator of circadian rhythms in animals (Karasek and Winczyk, 2006), and growing evidence suggests it also plays a role in the temporal regulation of physiological processes in plants (Fan et al., 2018). In horticultural crops, where developmental timing and flowering synchronization are critical for yield and quality, melatonin may contribute to the coordination of circadian and photoperiodic responses, although this field remains underexplored.
In plants, circadian rhythms regulate processes such as stomatal opening, photosynthetic activity, hormone synthesis and flowering time (Kim et al., 2017). Melatonin levels in plant tissues exhibit diurnal oscillations, with higher concentrations typically detected during the dark phase, suggesting endogenous circadian control of its biosynthesis (Mannino et al., 2021). This rhythmicity aligns with the expression patterns of melatonin biosynthetic genes such as TDC, SNAT and ASMT, which are influenced by light cues and the internal clock (Kumari et al., 2023). Melatonin has also been shown to influence circadian rhythms by modulating the expression of core clock genes and enhancing rhythm amplitude, as observed in Arabidopsis (Huang et al., 2012) and grapevine (Vitis vinifera L.) (Boccalandro et al., 2011). However, research on the circadian regulation of melatonin in horticultural crops remains extremely limited, and further studies are needed to elucidate its physiological relevance in these species.
Photoperiodism, the response of organisms to changes in day length, is a major determinant of flowering time in many horticultural species (Adams and Langton, 2005). While direct studies in horticultural crops are limited, similar mechanisms have been observed in other species. For instance, in apple trees (Malus domestica L.), melatonin levels fluctuate in response to seasonal light signals, and exogenous melatonin application has been found to delay flowering (Zhang et al., 2019). This suggests that melatonin may serve as a signal integrating environmental light cues with reproductive development across different plant species.
Additionally, melatonin’s photoprotective properties, including its ability to modulate chloroplast redox status and enhance light use efficiency, may help synchronize photosynthetic rhythms with environmental light regimes (Kumari et al., 2023). This contributes not only to improved stress resilience but also to optimized growth and resource use efficiency.
Overall, while the role of melatonin in circadian and photoperiodic regulation in horticultural crops is still an emerging field, preliminary findings indicate its potential involvement in aligning internal physiological rhythms with environmental cycles (Kumari et al., 2023). Further research in this area could reveal novel strategies to manipulate flowering time, optimize growth cycles, and improve productivity in diverse horticultural species.
MELATONIN AND ABIOTIC STRESS TOLERANCE IN HORTICULTURAL CROPS: MECHANISMS AND AGRONOMIC APPLICATIONS
Abiotic stresses such as drought, salinity, and temperature extremes represent major constraints to horticultural crop productivity and quality (Rouphael and Colla, 2020). These adverse conditions are becoming more frequent and severe due to climate change and environmental pressures, challenging conventional production systems and threatening food security (Abou-Hussein, 2012).
In recent years, growing interest has been directed toward eco-friendly and efficient strategies to enhance plant stress resilience. Among these, melatonin has emerged as a multifunctional molecule with strong potential to mitigate the effects of environmental stressors (Janas and Posmyk, 2013). While its physiological roles in plants have been broadly characterized, its specific applications in stress-prone horticultural systems require further synthesis.
This chapter explores the protective roles of melatonin in response to the main abiotic stresses in horticultural crops. For each type of stress, we examine its agronomic relevance, describe the known mechanisms of melatonin-induced tolerance, and highlight key examples from experimental and applied studies. We conclude with an overview of current and potential agronomic applications of melatonin within integrated crop management.
Drought stress
Drought is one of the most critical abiotic stress factors affecting agricultural production worldwide (Borgohain et al., 2019). Water scarcity severely disrupts plant physiological processes, including photosynthesis, nutrient transport, and cellular homeostasis, ultimately resulting in reduced growth, lower yields, and poor product quality (Seleiman et al., 2021). In horticultural crops, which often possess shallow root systems and high-water demands, drought stress can cause rapid and irreversible damage, particularly during reproductive stages (Rao et al., 2016).
Melatonin has been shown to play a significant role in enhancing drought tolerance through multiple and coordinated mechanisms. One of its most prominent actions is the modulation of the antioxidant defense system (Oloumi, 2022). Under drought conditions, ROS levels increase sharply, causing oxidative damage to lipids, proteins, and nucleic acids (Choudhury et al., 2017). Melatonin directly scavenges ROS and also upregulates antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and peroxidases, thereby maintaining cellular redox balance (Zhang and Zhang, 2014).
Beyond its antioxidant role, melatonin contributes to drought tolerance by regulating stomatal behavior and water use efficiency, in part through its interaction with abscisic acid (ABA), a key drought-responsive phytohormone. Melatonin modulates ABA biosynthesis and signaling pathways, thereby fine-tuning stomatal responses under water-deficient conditions (Muhammad et al., 2024). Additionally, it influences the expression of aquaporins and stress-related transcription factors, enhancing root water uptake and reducing transpiration (Sharma and Zheng, 2019).
Numerous studies have confirmed the beneficial effects of melatonin application in drought-stressed horticultural crops. In tomato, exogenous melatonin improved relative water content, chlorophyll concentration, and antioxidant enzyme activity, leading to better growth and yield under limited irrigation (Huang et al., 2023). In cucumber, melatonin pretreatment enhanced root development and delayed leaf wilting (Zhang et al., 2013). Similarly, in pepper (Capsicum annuum L.) and strawberry, melatonin treatments mitigated oxidative stress and preserved photosynthetic- efficiency during drought episodes (Kaya and Shabala, 2023; Khan et al., 2023).
These findings demonstrate that melatonin acts as a versatile regulator of drought tolerance in horticultural species, offering promising avenues for sustainable crop production under water-limiting conditions.
Salt stress
Salinity is a major abiotic constraint that limits the productivity and quality of crops across diverse agroecological zones (Safdar et al., 2019). While it is particularly prevalent in arid and semi-arid areas due to limited rainfall and saline irrigation, salinization also arises in other environments as a result of poor drainage, excessive fertilizer use, and soil degradation (Yadav et al., 2011).
High concentrations of sodium (Na⁺) and chloride (Cl⁻) ions disrupt cellular homeostasis by interfering with ion balance and water relations in plant tissues. This ionic imbalance leads to osmotic stress, nutrient deficiencies, membrane destabilization, and excessive generation of ROS, ultimately impairing photosynthesis, growth and productivity (Pandit et al., 2024; Safdar et al., 2019). As a result, salinity poses a significant challenge for sustainable crop production in affected regions.
Exogenous application of melatonin has been shown to significantly enhance salt stress tolerance in a wide range of horticultural crops. Its protective effects are attributed to its ability to modulate ion homeostasis, improve osmotic adjustment, reduce oxidative stress, and interact with hormonal pathways (Zhan et al., 2019). In tomato plants, melatonin treatment improved K⁺/Na⁺ ratios by regulating ion transporters such as SlSOS1, SlNHX1, and SlHKT1, thereby maintaining ionic balance under salt stress (Liu et al., 2015). Additionally, melatonin enhanced the activity of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), reducing ROS accumulation and lipid peroxidation in salt-stressed tissues.
In lettuce, melatonin alleviated salt-induced growth inhibition by enhancing the activity of antioxidant enzymes and limiting the accumulation of Na⁺ and Cl⁻ ions, probably through the modulation of stress-responsive gene expression (El-Bauome et al., 2024). Similarly, in melon (Cucumis melo L.), foliar application of melatonin combined with Ca²⁺ improved photosynthetic efficiency, water use efficiency, and ion homeostasis under saline conditions by decreasing Na⁺ levels and increasing Ca²⁺ accumulation in both leaves and roots (Wu et al., 2019). In this species, seed priming with melatonin has also been shown to enhance germination rates under salt stress, indicating its effectiveness at early developmental stages (Castañares and Bouzo, 2019).
Melatonin’s role in hormonal crosstalk under salinity has also been documented. It has been shown to interact with abscisic acid (ABA), gibberellins (GAs), and ethylene signaling pathways, contributing to fine-tuned stress responses. For instance, melatonin application downregulates ABA biosynthesis genes under moderate salt stress, preventing excessive stomatal closure and allowing continued gas exchange and growth.
Taken together, these findings support the use of melatonin as a promising biostimulant for mitigating salt stress in horticultural systems. Its multifaceted mode of action, involving ion transport regulation, antioxidant defense enhancement, osmotic adjustment, and hormonal interactions, provides a strong basis for its integration into sustainable crop management strategies.
Temperature stress
Temperature extremes, both high and low, pose significant threats to the productivity and quality of horticultural crops. Heat stress can disrupt photosynthesis, accelerate respiration, denature proteins and compromise reproductive development (Jagadish et al., 2021), while cold stress affects membrane fluidity, enzyme activity, and nutrient transport (Soualiou et al., 2022). As global climate change increases the frequency and intensity of thermal fluctuations, enhancing crop resilience to temperature stress has become a critical priority in horticultural systems (Devireddy et al., 2021).
Melatonin has been shown to enhance thermotolerance in plants by protecting the photosynthetic apparatus, reducing oxidative damage, and regulating the expression of heat-responsive genes (Hassan et al., 2022). In tomato, exogenous melatonin increased chlorophyll content and antioxidant enzyme activity, while upregulating genes encoding heat shock proteins (HSPs) (Xu et al., 2016). Similarly, in strawberry and celery (Apium graveolens L.), melatonin treatment enhanced thermotolerance by improving photosynthetic efficiency, reducing oxidative stress, and promoting the expression of heat-responsive genes, resulting in better seedling survival under high temperature conditions (Li et al., 2022; Manafi et al., 2022).
Cold stress tolerance is also enhanced by melatonin through mechanisms that include ROS scavenging, maintenance of membrane integrity, and regulation of cold-responsive pathways (Qari et al., 2022). In pepper, melatonin application under cold stress improved photosynthetic pigments, enzyme activities, and photosystem efficiency, resulting in better cold tolerance (Altaf et al., 2022). Similarly, in cucumber, foliar application of melatonin under low temperature and high humidity conditions enhanced photosynthetic efficiency, stimulated the antioxidant defense system and improved nutrient uptake, contributing to increased tolerance to cold stress (Amin et al., 2022).
Overall, the application of melatonin represents a promising approach to mitigate the detrimental effects of both heat and cold stress in horticultural crops, supporting plant survival and productivity under increasingly variable climatic conditions.
Figure 2 illustrates how melatonin enhances abiotic stress tolerance in horticultural crops, complementing the findings in table 1 and the earlier case studies.
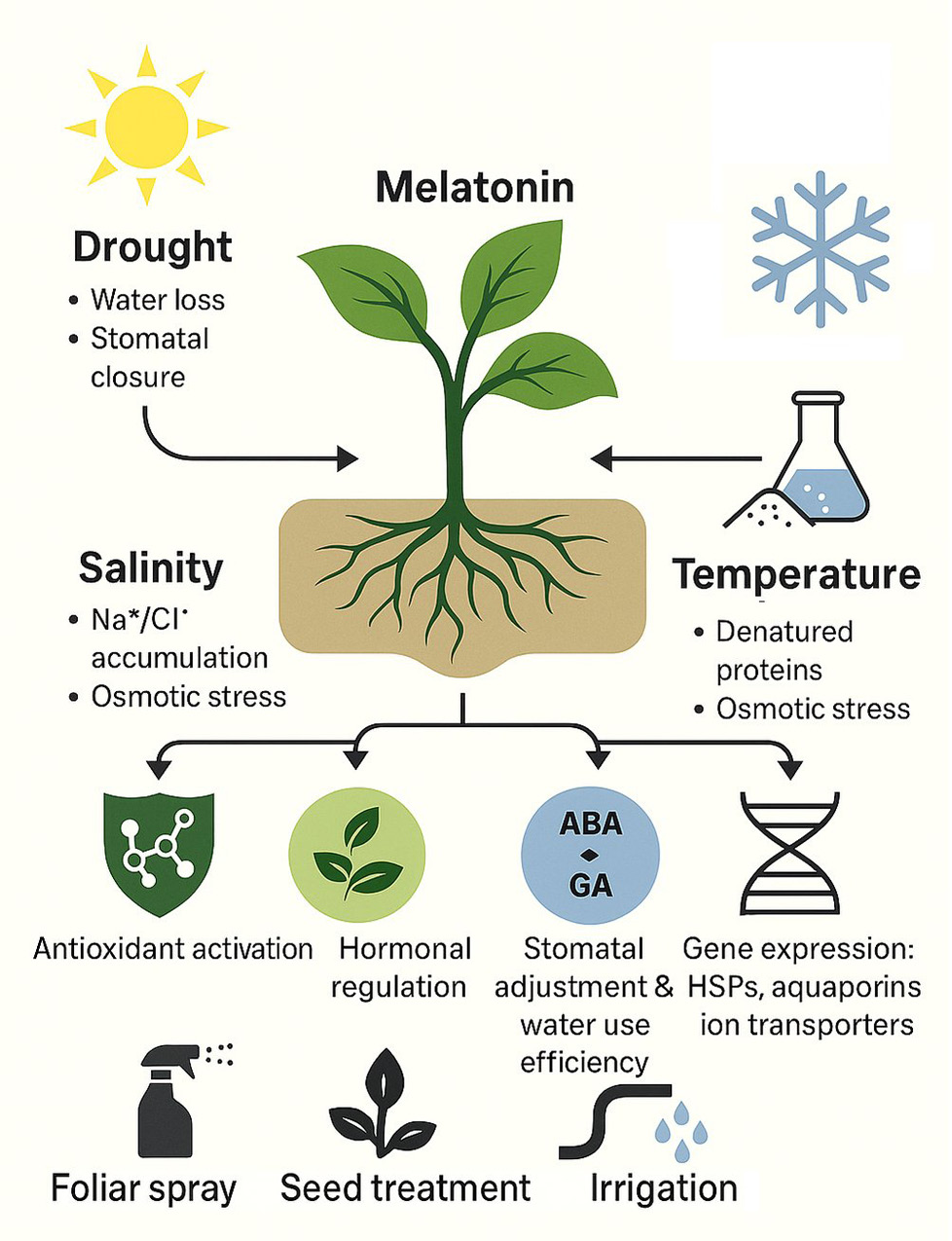
Figure 2.
Melatonin-mediated abiotic stress tolerance in horticultural crops. The figure summarizes how melatonin mitigates drought, salinity, and temperature stress through antioxidant activation, hormonal modulation, and stress-responsive gene regulation, alongside common agronomic application methods.
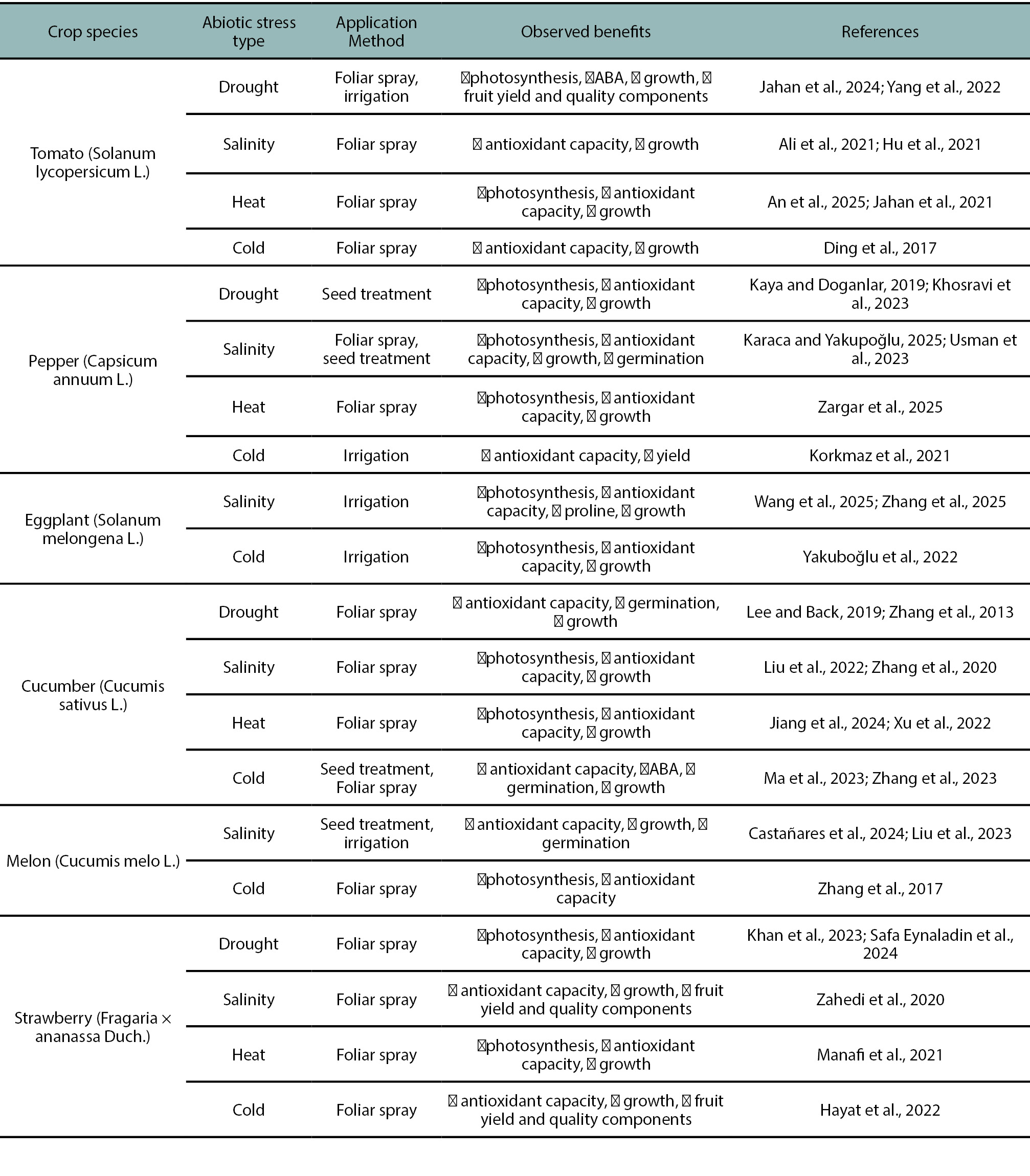
Table 1.
Effects of exogenous melatonin application on abiotic stress tolerance in some horticultural crop.
FUTURE DIRECTIONS AND CHALLENGES IN THE USE OF MELATONIN FOR SUSTAINABLE HORTICULTURE
Knowledge gaps and research needs
Despite substantial progress in understanding the roles of melatonin in horticultural crops, several critical knowledge gaps remain. The precise biosynthetic pathways and regulation of endogenous melatonin under different environmental conditions are not fully elucidated, limiting the ability to manipulate its levels effectively. In addition, the full spectrum of melatonin receptors and the downstream signaling pathways involved in transducing its effects have yet to be comprehensively identified and characterized (Kumari et al., 2023). This lack of molecular insight constrains our understanding of how melatonin elicits specific physiological responses under stress conditions.
Moreover, the complex interactions between melatonin and plant-associated microbiomes, as well as its combined effects with other biostimulants and phytohormones, remain poorly understood (Arnao and Hernández-Ruiz, 2019). Addressing these research needs will be crucial to fully exploit melatonin’s potential in sustainable horticulture.
Technological and agronomic challenges in melatonin application
Although melatonin has shown great promise as a biostimulant in horticulture, several practical challenges hinder its widespread application. A key limitation is the lack of standardized protocols for dosage, application methods (e.g., foliar spray, seed priming), and timing, which often leads to inconsistent outcomes across different species and environmental conditions (Arnao and Hernández-Ruiz, 2019). Moreover, melatonin is not yet officially recognized as a biostimulant or plant growth regulator in most regulatory frameworks, restricting its commercialization and broader adoption (Arnao and Hernández-Ruiz, 2019).
Additional barriers include its chemical instability under field conditions —particularly its sensitivity to light and temperature— which can reduce its effectiveness. To overcome this, advanced delivery systems such as nano-formulations and encapsulation are being developed to improve its stability and ensure controlled release (Mukherjee et al., 2024). At the same time, the limited availability and high cost of commercial melatonin products tailored for agriculture represent a further obstacle to large-scale implementation (Arnao and Hernández-Ruiz, 2019).
In summary, advancing melatonin use in horticulture will require optimizing application strategies, improving formulation technologies, and establishing clear regulatory frameworks to ensure efficacy, affordability, and environmental safety.
Melatonin has emerged as a multifaceted regulator in horticultural crops, playing critical roles in growth, development, and especially in enhancing tolerance to various abiotic stresses. Its antioxidant properties, interaction with phytohormones, and modulation of physiological and molecular responses position melatonin as a promising tool for sustainable horticulture. However, despite significant advances, key gaps remain in our understanding of its biosynthesis, signaling pathways and interactions with plant microbiomes, which limit the full exploitation of its potential.
The practical application of melatonin in the field is challenged by the lack of standardized protocols, formulation stability issues and regulatory hurdles. Addressing these challenges through multidisciplinary research and development of tailored delivery systems will be essential for its effective integration into crop management practices.
Looking forward, future studies should focus on elucidating the molecular mechanisms underlying melatonin’s actions, optimizing application strategies for diverse horticultural species, and evaluating long-term environmental impacts. Such efforts will contribute to unlocking the full benefits of melatonin, enhancing crop resilience and productivity in the face of increasing environmental stresses.
In conclusion, melatonin represents a valuable biostimulant with great promise for advancing sustainable horticulture, provided that current knowledge gaps and practical challenges.
redalyc-journal-id: 864
Figure 1.
Melatonin biosynthesis pathway in plants (adapted from Yan et al., 2023).
Figure 2.
Melatonin-mediated abiotic stress tolerance in horticultural crops. The figure summarizes how melatonin mitigates drought, salinity, and temperature stress through antioxidant activation, hormonal modulation, and stress-responsive gene regulation, alongside common agronomic application methods.
Table 1.
Effects of exogenous melatonin application on abiotic stress tolerance in some horticultural crop.
